
研究活動 Research
論文紹介 Highlights
Highlights in 2025

論文
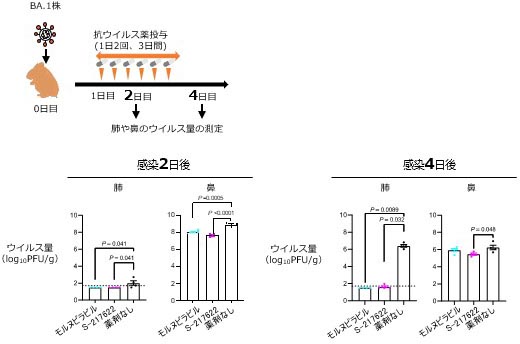
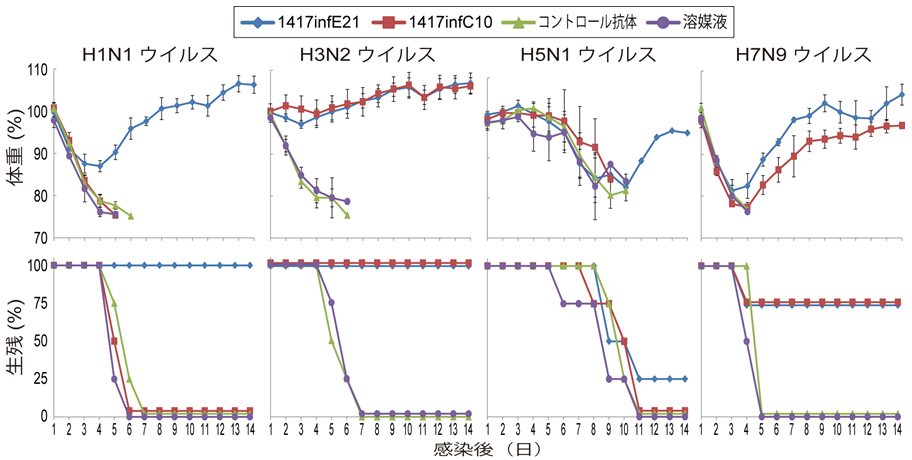
マウスモデルにおける牛由来高病原性H5N1鳥インフルエンザウイルスに対するバロキサビルの効果
Efficacy of baloxavir marboxil against bovine H5N1 virus in mice
20 June 2025

論文
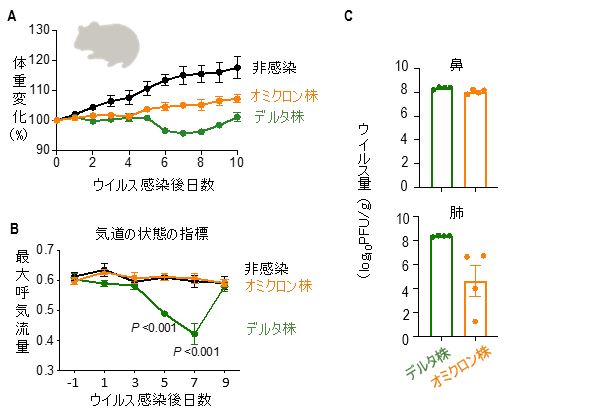
新型コロナウイルスXBB.1.5型RBDを発現するmRNAワクチンの抗原性の異なるJN.1株に対する有効性の検証
An mRNA vaccine encoding the SARS-CoV-2 Omicron XBB.1.5 receptor-binding domain protects mice from the JN.1 variant
6 June 2025

論文
動物モデルを用いた 新型コロナウイルス 半生ウイルスワクチンプラットフォームの確立
SARS-CoV-2 virus lacking the envelope and membrane open-reading frames as a vaccine platform
14 May 2025

論文
乳牛の乳房・乳頭組織におけるウシ由来H5N1高病原性 鳥インフルエンザウイルスの増殖性状を解明
Highly pathogenic avian H5N1 influenza A virus replication in ex vivo cultures of bovine mammary gland and teat tissues
→ Emerging Microbes & Infections
15 January 2025

News
NEJM Outbreaks Update — H5N1
鳥インフルエンザが鶏、牛、猫、そして人にどのような影響を及ぼしているのか?NEJM編集長 Eric Rubin、副編集長 Lindsey Badenと共に、河岡義裕教授が最新の状況を解説します。
The New England Journal of Medicine 392(12):e34
5 March 2025

論文
免疫細胞が引き起こすCOVID-19の重症化機序を解明 ― 肺血管での異常な接着現象に着目 ―
Neutrophil adhesion to vessel walls impairs pulmonary circulation in COVID-19 pathology
14 January 2025
Highlights in 2024

論文
高病原性鳥インフルエンザH5HAのLNP-mRNAワクチンは牛由来H5N1 ウイルスに対して防御効果を示す
Protective effects of an mRNA vaccine candidate encoding H5HA clade 2.3.4.4b against the newly emerged dairy cattle H5N1 virus
30 October 2024

論文
ヒトから分離された牛由来高病原性H5N1鳥インフルエンザウイルスの病原性と感染伝播性
A human isolate of bovine H5N1 is transmissible and lethal in animal models
28 October 2024

論文
牛由来高病原性H5N1鳥インフルエンザウイルスの マウスおよびフェレットにおける病原性と伝播性
Pathogenicity and transmissibility of bovine H5N1 influenza virus
8 July 2024

論文
牛乳から検出された高病原性H5N1鳥インフルエンザウイルスの熱不活性化とマウスへの感染性
Cow’s Milk Containing Avian Influenza A(H5N1) Virus — Heat Inactivation and Infectivity in Mice
→ New England Journal of Medicine
24 May 2024
Highlights in 2023
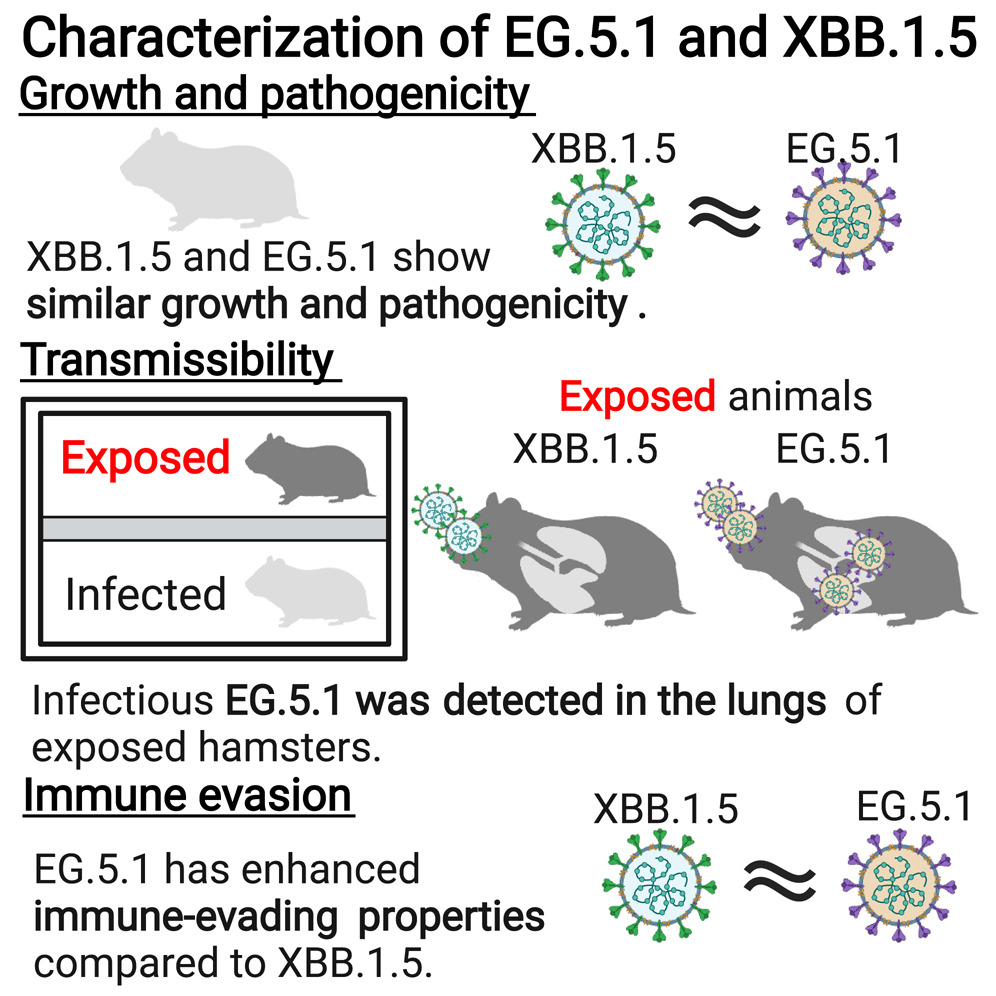
論文
新型コロナウイルス・オミクロン株EG.5.1系統は、 ハムスターで初期のオミクロン系統よりも伝播しやすい
Characterization of a SARS-CoV-2 EG.5.1 clinical isolate in vitro and in vivo
15 December 2023
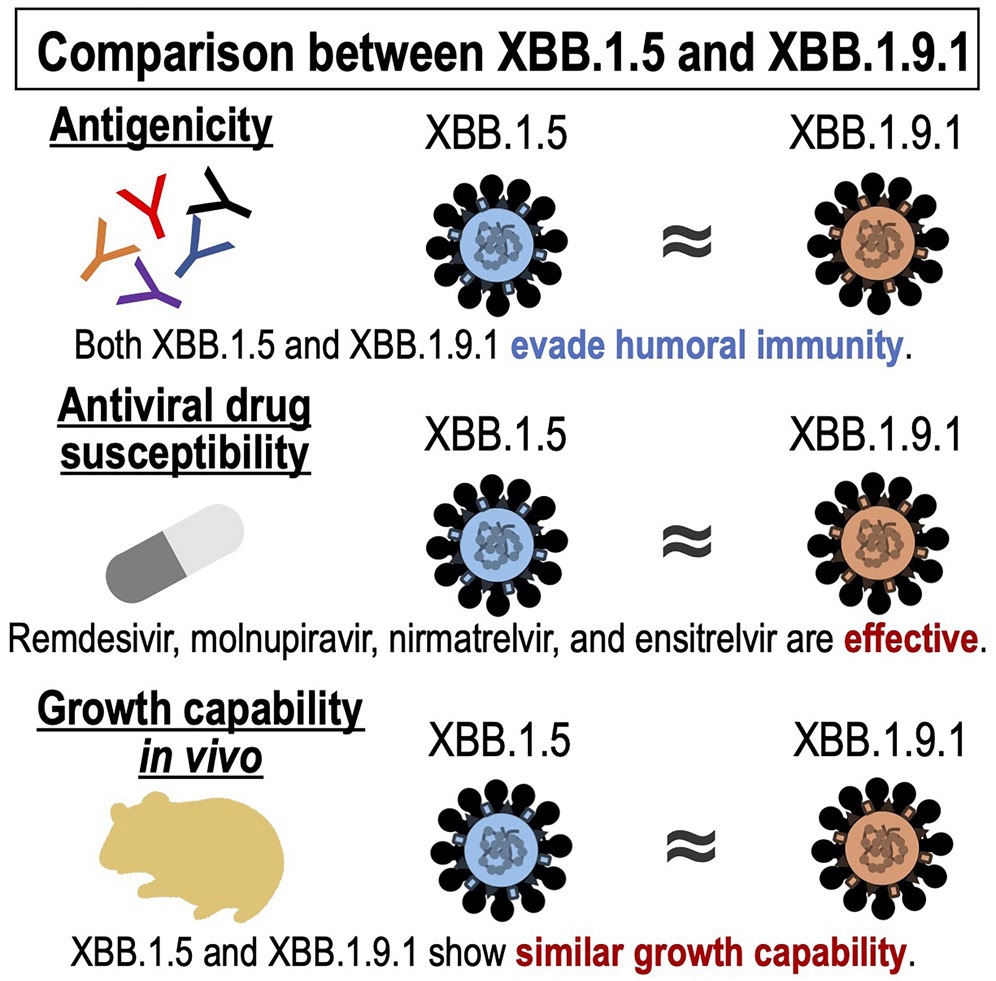
論文
新型コロナウイルス・オミクロン株のXBB.1.9.1系統はXBB.1.5系統と類似の性状を持つ
Antiviral efficacy against and replicative fitness of an XBB.1.9.1 clinical isolate
4 October 2023
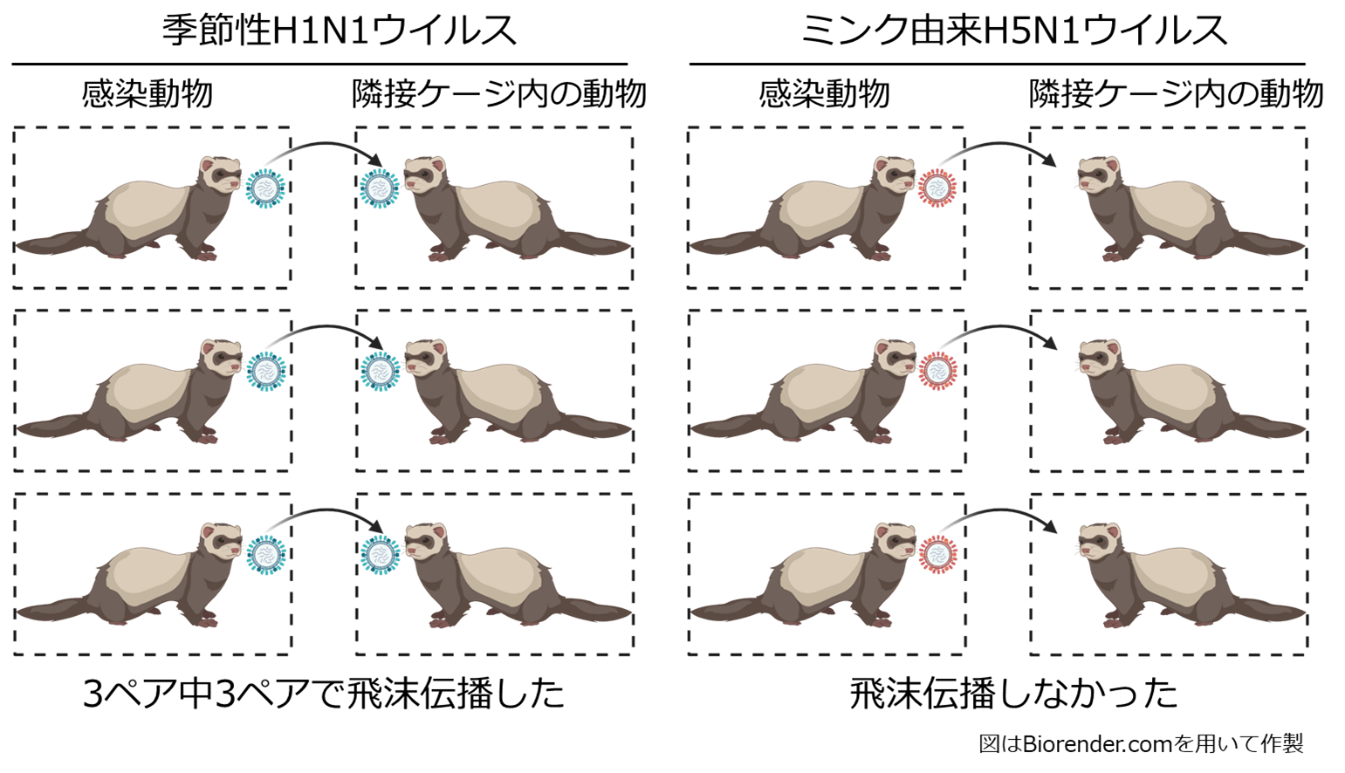
論文
ミンク由来高病原性H5N1鳥インフルエンザウイルスの性状解析
Characterization of highly pathogenic clade 2.3.4.4b H5N1 mink influenza viruses
7 October 2023
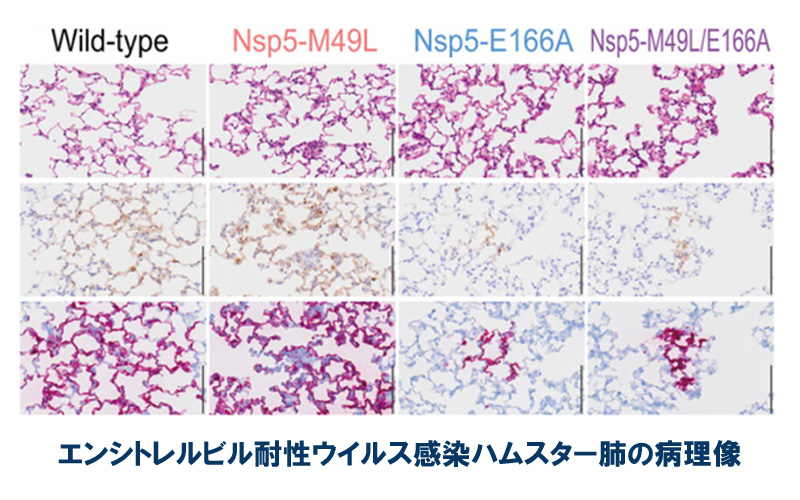
論文
ゾコーバ耐性新型コロナウイルスの性状解析
In vitro and in vivo characterization of SARS-CoV-2 resistance to ensitrelvir
25 July 2023
論文
インフルエンザウイルスHAの変異し難い部位をねらう: ワクチン抗原のデザイン
ユニバーサルワクチンの新規ストラテジー
Influenza H3 hemagglutinin vaccine with scrambled immunodominant epitopes elicits antibodies directed toward immunosubdominant head epitopes
19 July 2023
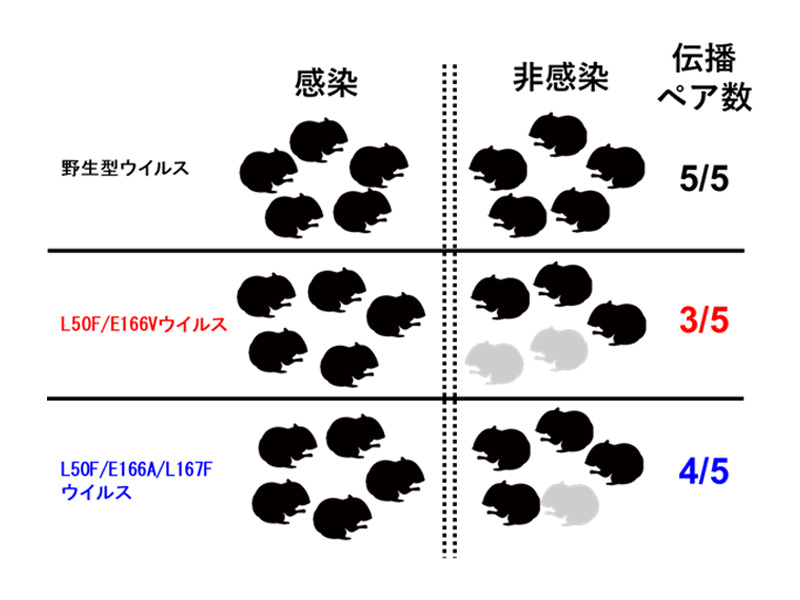
論文
ニルマトレルビル耐性新型コロナウイルスの性状解析
In vitro and in vivo characterization of SARS-CoV-2 strains resistant to nirmatrelvir
4 July 2023
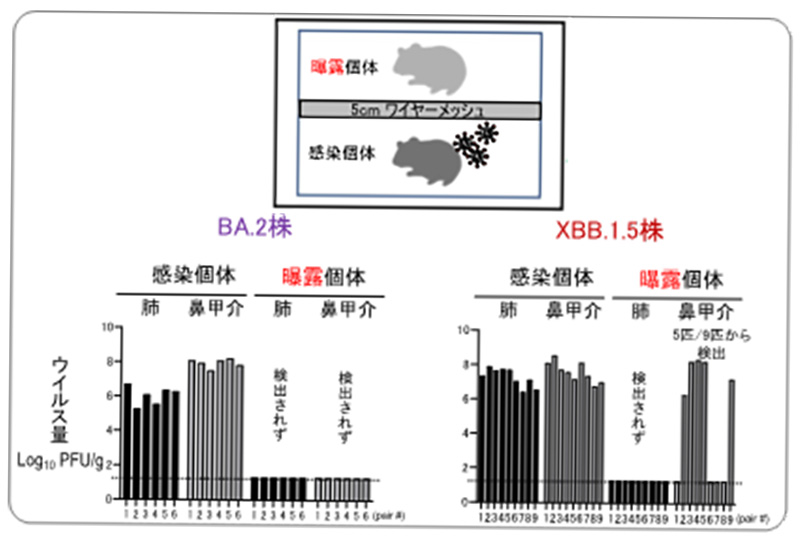
論文
新型コロナウイルス・オミクロン株XBB.1.5系統は、ハムスターで初期のオミクロン系統よりも伝播しやすい
Transmission and re-infection of Omicron variant XBB.1.5 in hamsters
21 June 2023
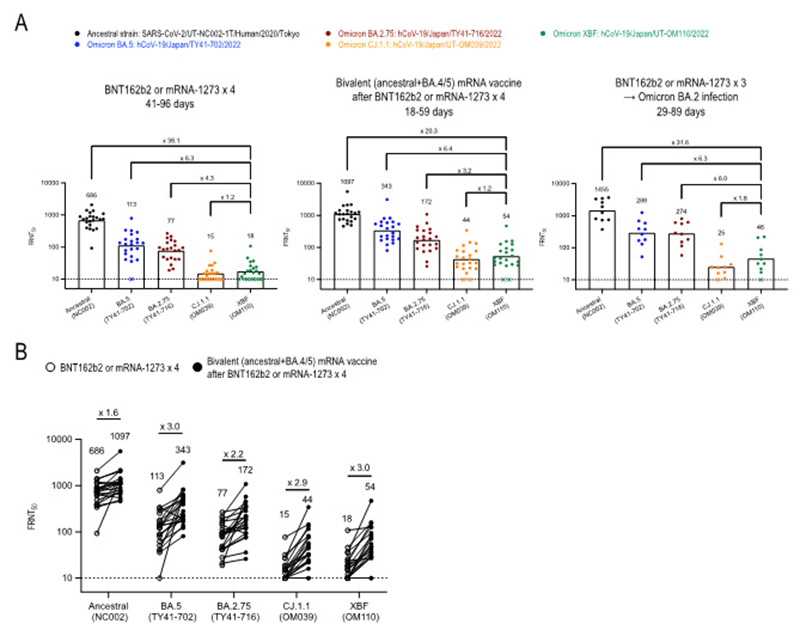
論文
BA.4/5株対応2 価mRNAワクチン接種で、新型コロナウイルス・オミクロン株XBF系統に対する中和活性は増強される
Efficacy of antivirals and mRNA vaccination against an XBF clinical isolate
→ Lancet Regional Health - Western Pacific
11 May 2023
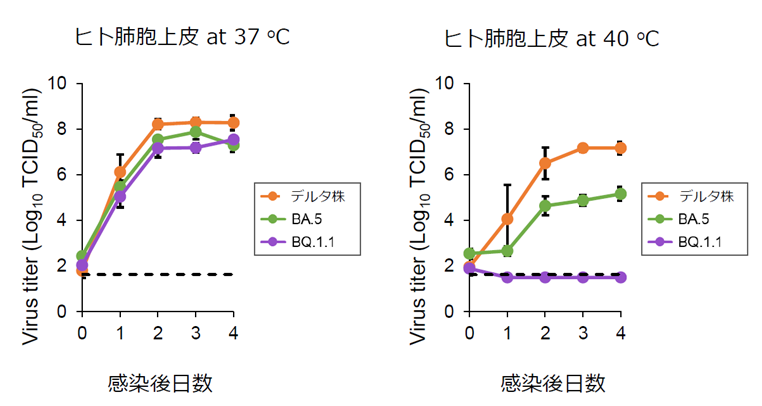
論文
新型コロナウイルス・オミクロン株のBA.5系統およびBQ.1.1系統が、高温で増殖しづらいことを解明
Replicative capacity of SARS-CoV-2 omicron variants BA.5 and BQ.1.1 at elevated temperatures
24 April 2023
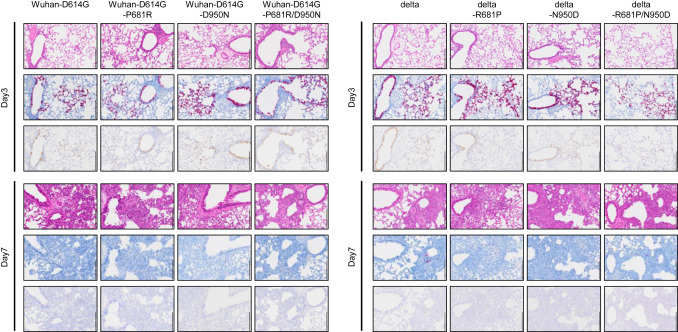
論文
新型コロナウイルス・デルタ変異株の Spike-P681RおよびD950N変異の機能解析
In SARS-CoV-2 delta variants, Spike-P681R and D950N promote membrane fusion, Spike-P681R enhances spike cleavage, but neither substitution affects pathogenicity in hamsters
10 April 2023
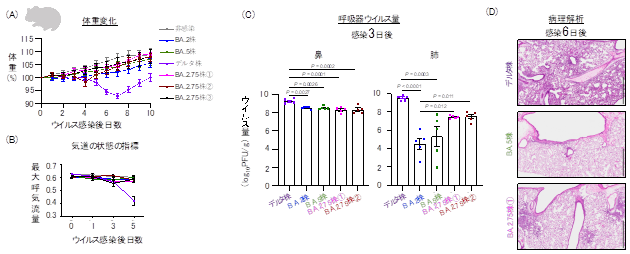
論文
新型コロナウイルス・オミクロン変異株のBA.2.75系統の性状解析
Characterization of SARS-CoV-2 Omicron BA.2.75 clinical isolates
23 March 2023
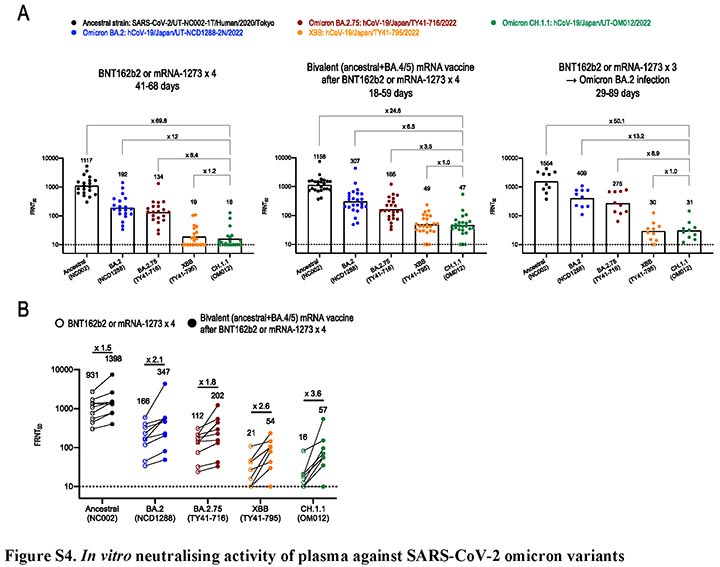
新型コロナウイルス・オミクロン株のCH.1.1系統に対する 治療薬と2価ワクチンの効果を検証
Efficacy of Antiviral Agents and Bivalent mRNA Vaccines against a CH.1.1 isolate
7 March 2023
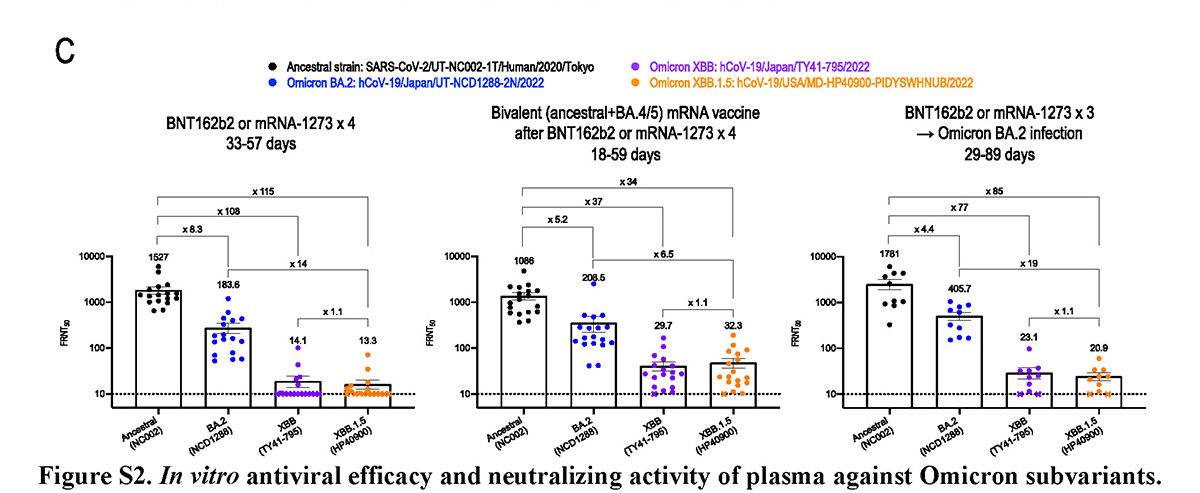
新型コロナウイルス・オミクロン株のXBB.1.5系統に対する 治療薬とワクチンの効果を検証
Antiviral and bivalent vaccine efficacy against an omicron XBB.1.5 isolate
8 February 2023
Highlights in 2022
新型コロナウイルス感染症とインフルエンザが同時流行しているかを検証
Are twindemics occurring?
→ Influenza and Other Respiratory Viruses
25 December 2022
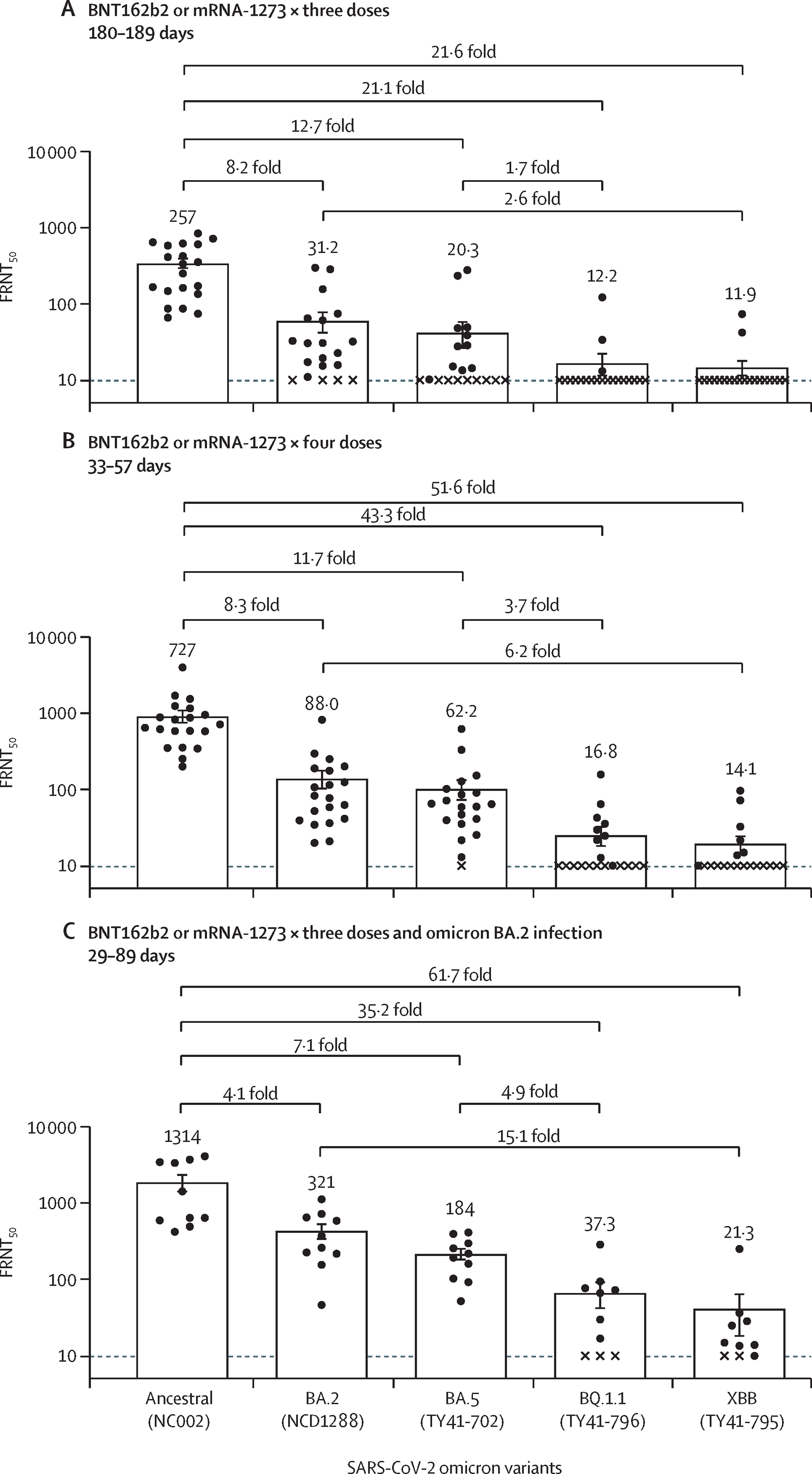
ワクチン被接種者あるいはブレイクスルー感染者血漿の、 臨床検体から分離した新型コロナウイルス・オミクロン株の BQ.1.1系統とXBB系統に対する反応性を検証
Humoral immune evasion of the omicron subvariants BQ.1.1 and XBB
7 December 2022
 写真: 新型コロナウイルス・オミクロン変異株
写真: 新型コロナウイルス・オミクロン変異株BA.5系統の走査型電子顕微鏡写真
患者から分離した新型コロナウイルス・オミクロン変異株のBA.4系統ならびにBA.5系統の性状解明
Characterization of SARS-CoV-2 Omicron BA.4 and BA.5 isolates in rodents
2 November 2022
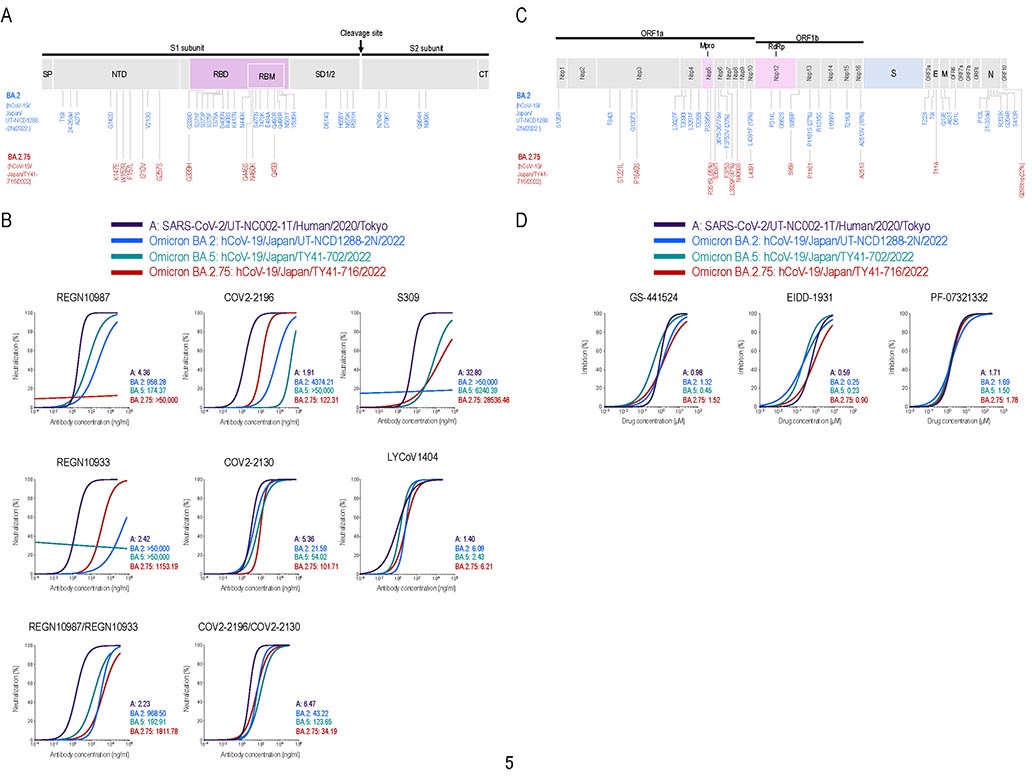
臨床検体から分離した新型コロナウイルス・オミクロン株 BA.2.75系統に対する治療薬の効果を検証
Efficacy of Antiviral Agents against the Omicron Subvariant BA.2.75
→ New England Journal of Medicine
7 September 2022
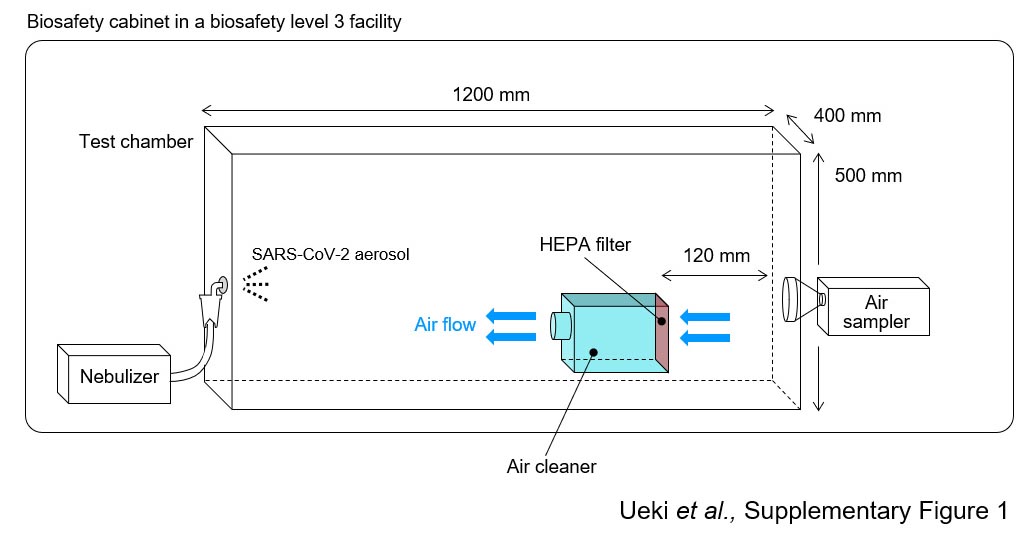
HEPAフィルターによる エアロゾル中の感染性新型コロナウイルスの除去効果
Effectiveness of HEPA filters at removing infectious SARS-CoV-2 from the air
10 August 2022
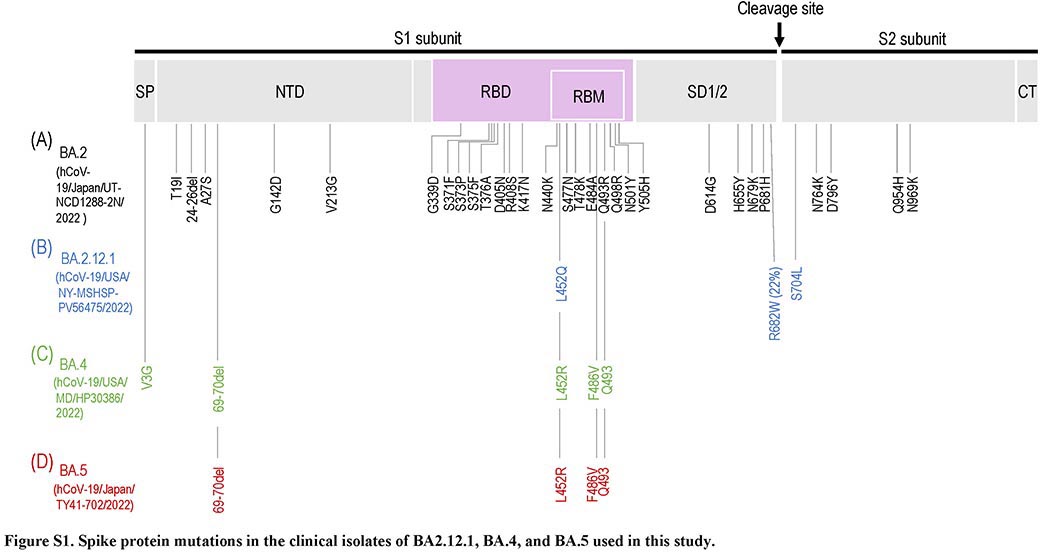
臨床検体から分離した新型コロナウイルス・オミクロン株 BA.2.12.1、BA.4、BA.5系統に対する治療薬の効果を検証
Efficacy of Antibodies and Antiviral Drugs against Omicron BA.2.12.1, BA.4, and BA.5 Subvariants
→ New England Journal of Medicine
20 July 2022
新型コロナウイルス・オミクロン/BA.1変異株に対する 治療薬の効果の検証 — 抗ウイルス薬を投与した免疫不全動物において耐性ウイルスが出現するリスクは低い —
Therapeutic efficacy of monoclonal antibodies and antivirals against SARS-CoV-2 Omicron BA.1 in Syrian hamsters
15 June 2022
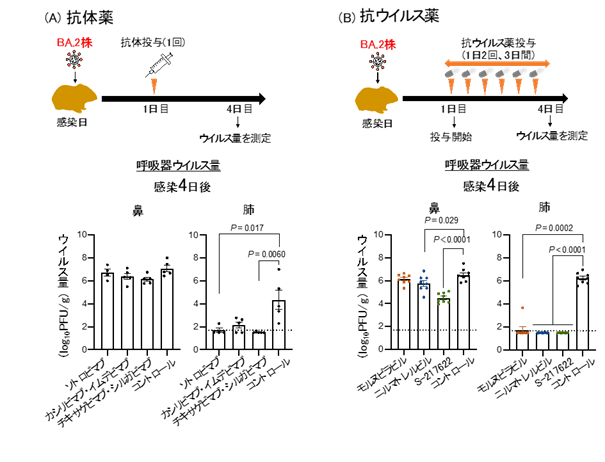
患者から分離された新型コロナウイルス・オミクロン/BA.2変異株の性状解明 — オミクロン/BA.1変異株と同程度の病原性であることが明らかに —
Characterization and antiviral susceptibility of SARS-CoV-2 Omicron/BA.2
16 May 2022
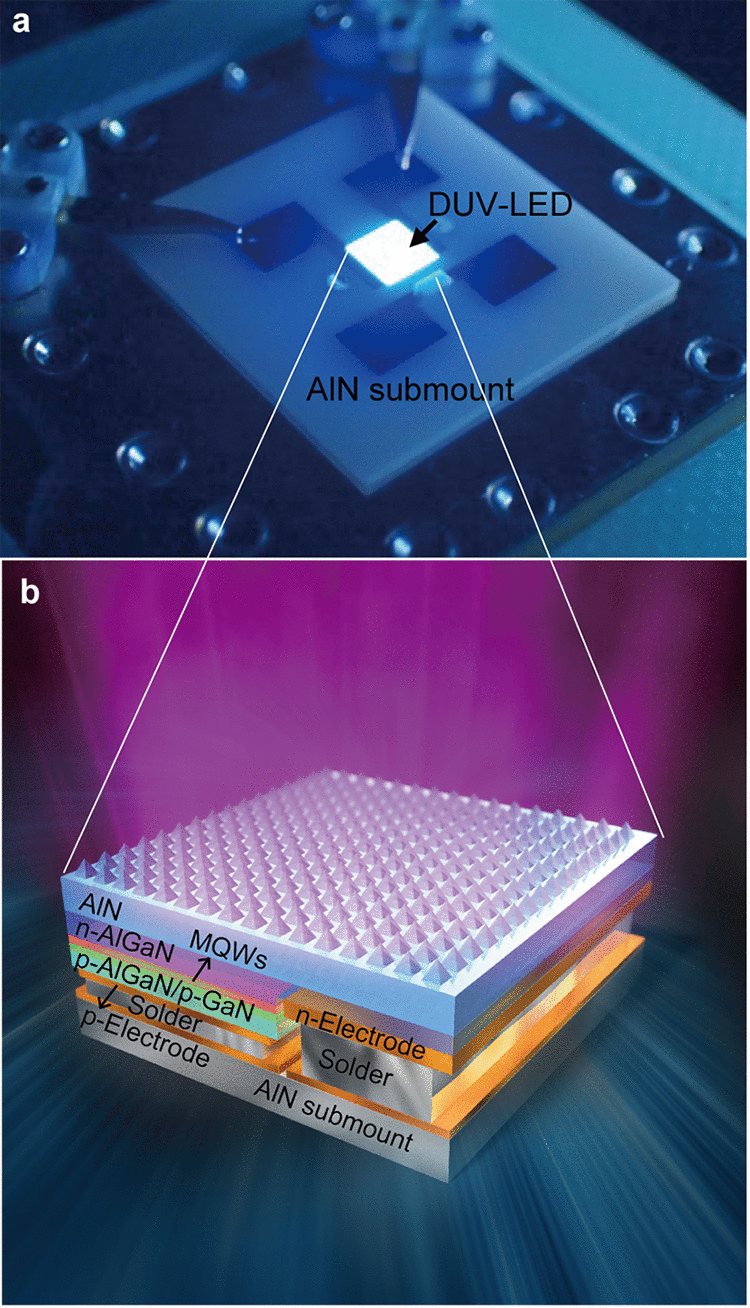
高出力深紫外LED(265nm帯)により エアロゾル中の新型コロナウイルスの高速不活性化に成功
A 265-Nanometer High-Power Deep-UV Light-Emitting Diode Rapidly Inactivates SARS-CoV-2 Aerosols
17 March 2022
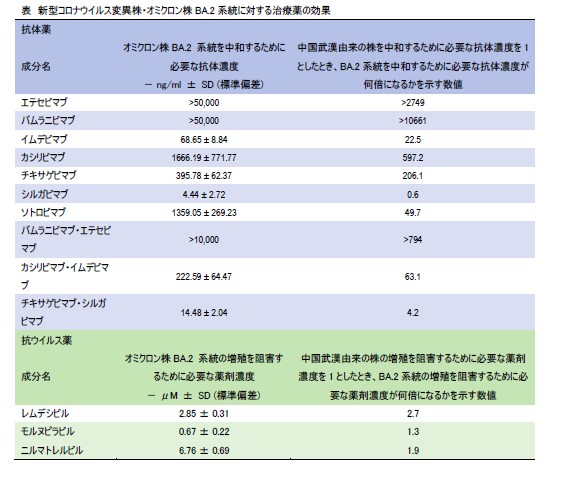
新型コロナウイルス変異株・オミクロン株BA.2系統に対する治療薬の効果を検証
Efficacy of Antiviral Agents against the SARS-CoV-2 Omicron Subvariant BA.2
→ New England Journal of Medicine
9 March 2022
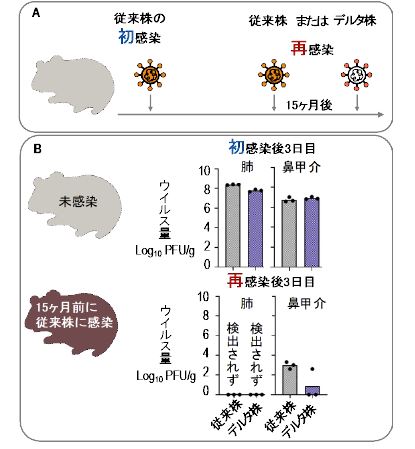
パンデミック初期のSARS-CoV-2株感染によって誘導された免疫は、 長期にわたって維持され、デルタ株による再感染を防御する
Long-term, infection-acquired immunity against the SARS-CoV-2 Delta variant in a hamster model
15 February 2022
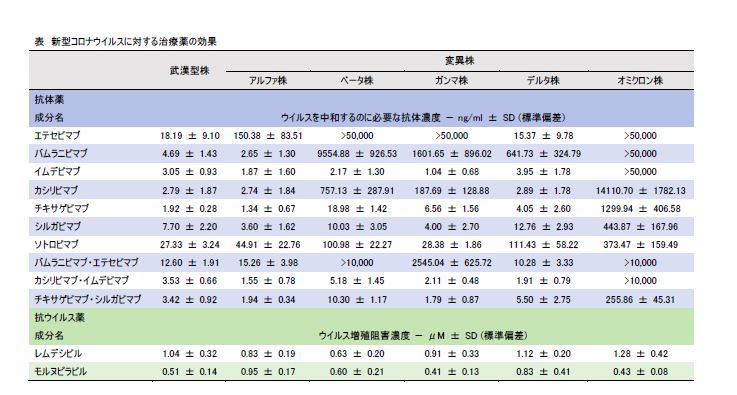
新型コロナウイルス変異株・オミクロン株に対する治療薬の効果を検証
Efficacy of Antibodies and Antiviral Drugs against Covid-19 Omicron Variant
→ New England Journal of Medicine
26 January 2022
Highlights in 2021
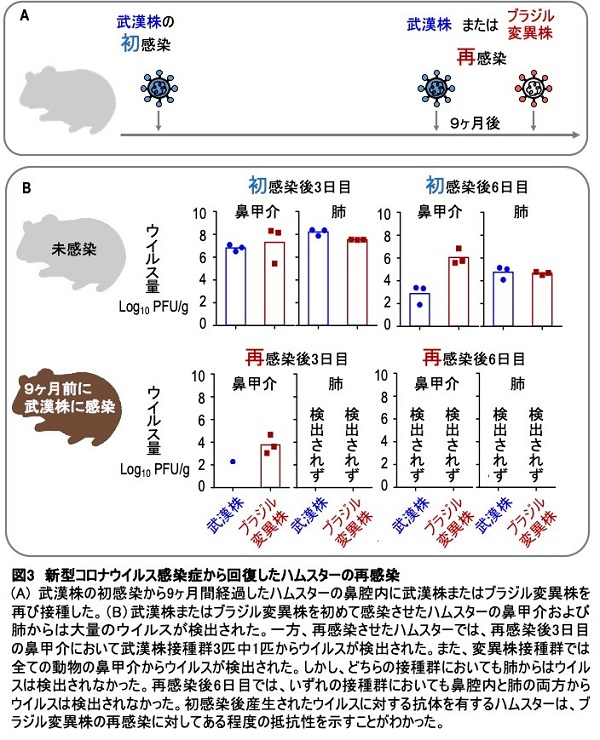
ブラジルからの帰国者から分離された新型コロナウイルス変異株の性状解明、 ならびに本変異株に対するワクチン効果の検証
Characterization of a new SARS-CoV-2 variant that emerged in Brazil
17 June 2021
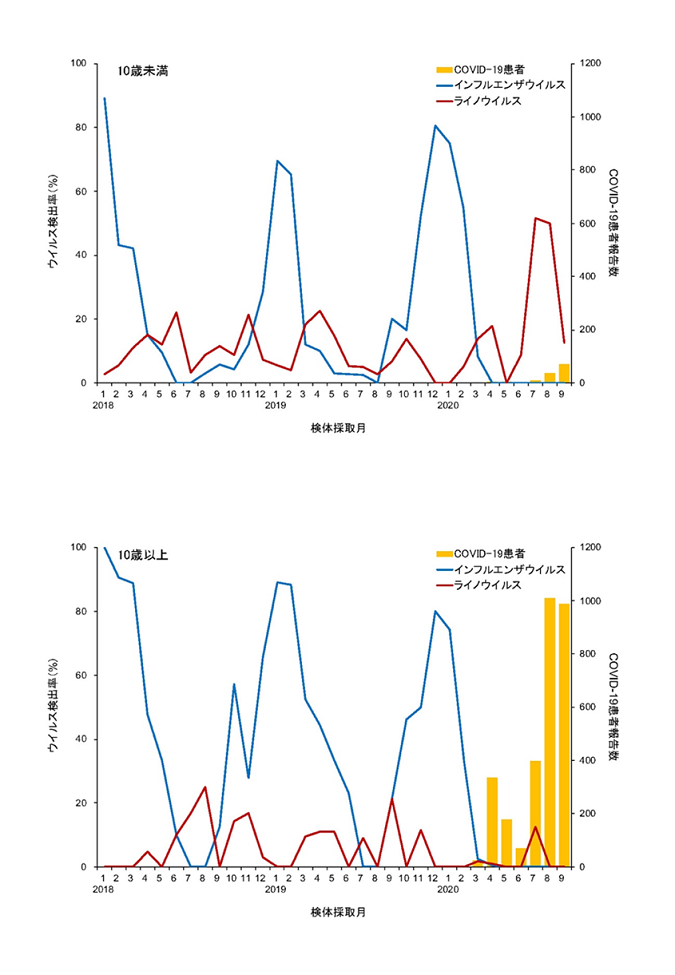 横浜市におけるインフルエンザ及びライノウイルス
横浜市におけるインフルエンザ及びライノウイルス検出率と新型コロナウイルス感染症(COVID-19)
患者報告数
新型コロナウイルス感染症流行下では 10歳未満の小児のライノウイルス感染リスクが上昇した
Increased risk of rhinovirus infection in children during the coronavirus disease-19 pandemic
→ Influenza and Other Respiratory Viruses
15 March 2021
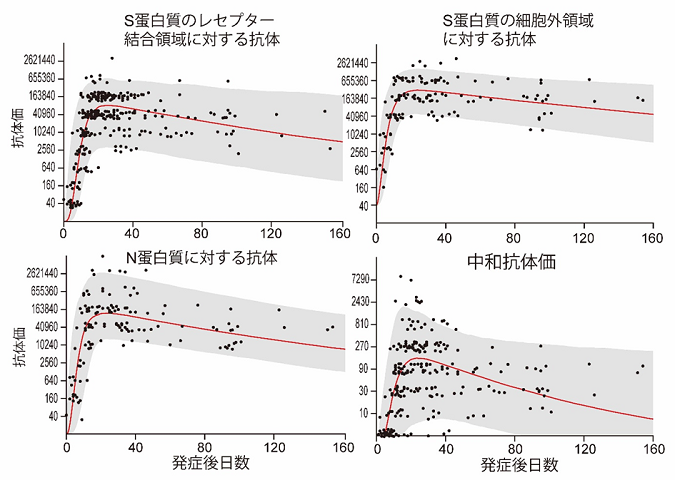 COVID-19患者におけるSARS-CoV-2に対する抗体価の変動
COVID-19患者におけるSARS-CoV-2に対する抗体価の変動
新型コロナウイルス感染時に獲得されたウイルスに対する抗体は 少なくとも発症後3~6か月間は維持される
Antibody titers against SARS-CoV-2 decline, but do not disappear for several months
12 February 2021
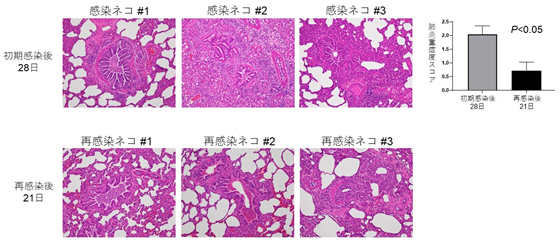
新型コロナウイルスに感染し回復したネコは無症状にもかかわらず 長期間、肺に炎症ダメージが残り、また一定期間は再感染しない
Protective Immunity and Persistent Lung Sequelae in Domestic Cats after SARS-CoV-2 Infection
→ Emerging Infectious Diseases
7 January 2021
Highlights in 2020
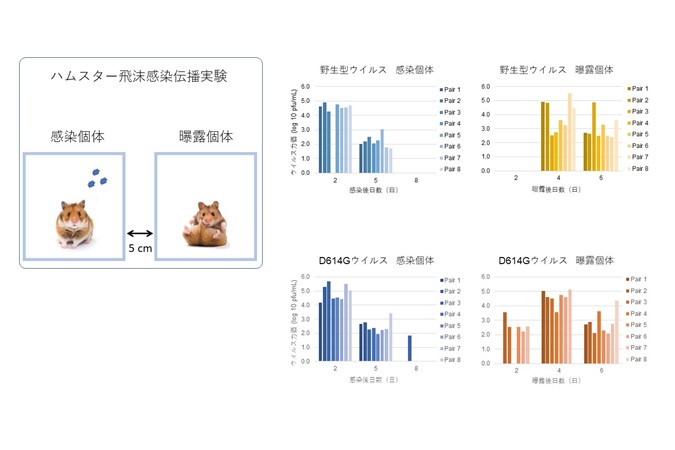 ハムスターにおけるSARS-CoV-2 野生型・
ハムスターにおけるSARS-CoV-2 野生型・D614Gウイルスの飛沫感染伝播実験
現在流行中のSARS-CoV-2 D614G変異株は、高い増殖効率と感染伝播力を示す
SARS-CoV-2 D614G Variant Exhibits Efficient Replication ex vivo and Transmission in vivo
12 November 2020
 BSL3施設内に設置したウイルス噴霧チャンバー
BSL3施設内に設置したウイルス噴霧チャンバー
新型コロナウイルスの空気伝播に対するマスクの防御効果
Effectiveness of face masks in preventing airborne transmission of SARS-CoV-2
21 October 2020
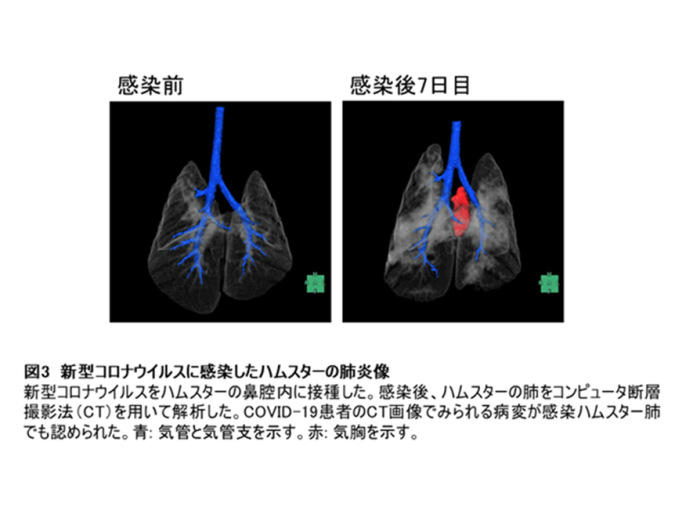
新型コロナウイルス感染症( COVID-19 )の病態解明 / 予防・治療法の開発 ハムスターの感染動物モデルとしての有用性
Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development
22 June 2020
 ネコを用いた新型コロナウイルス感染伝播実験
ネコを用いた新型コロナウイルス感染伝播実験
新型コロナウイルスはネコの間で感染伝播する
Transmission of SARS-CoV-2 in Domestic Cats
→ New England Journal of Medicine
13 May 2020
Highlights in 2019
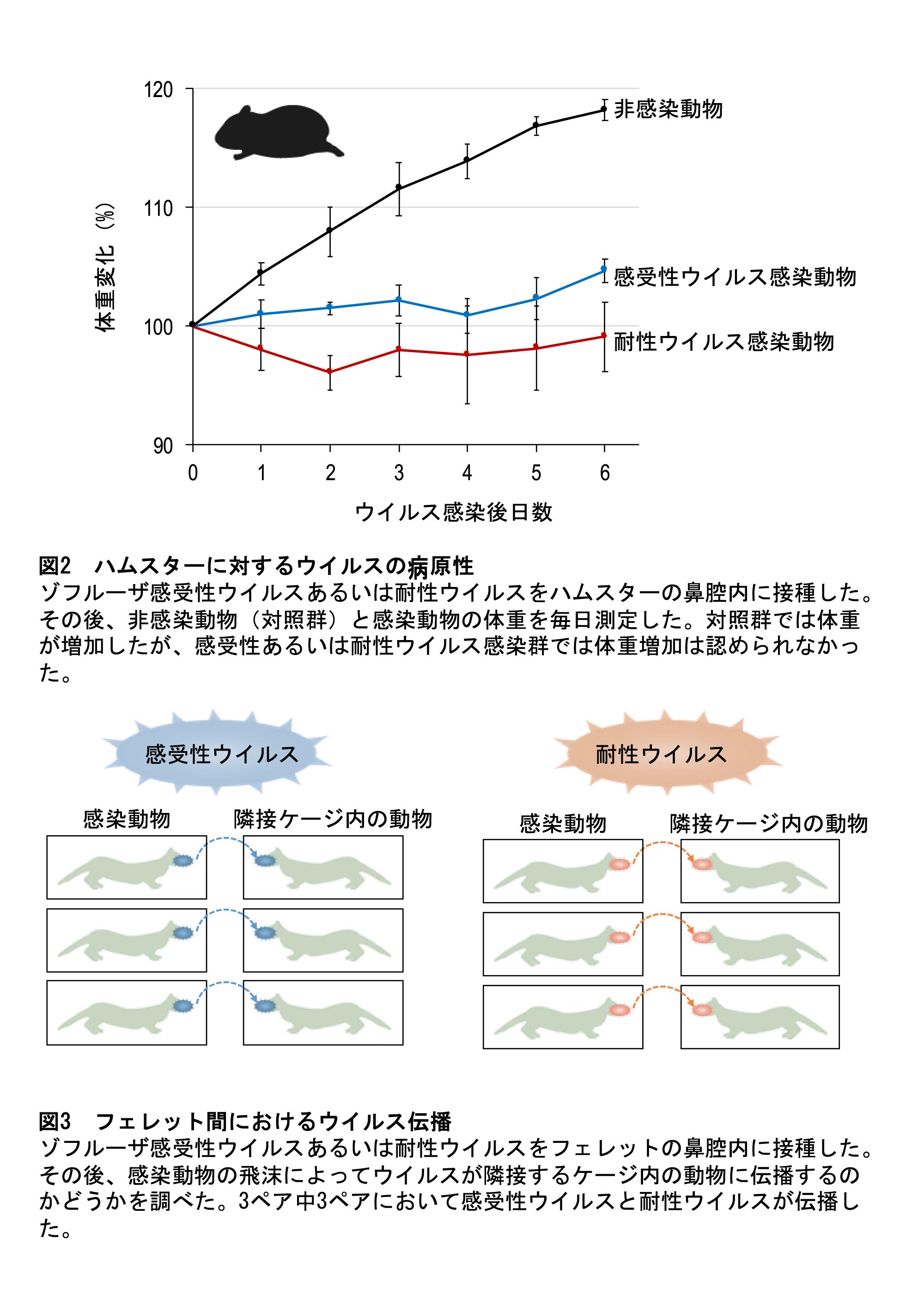
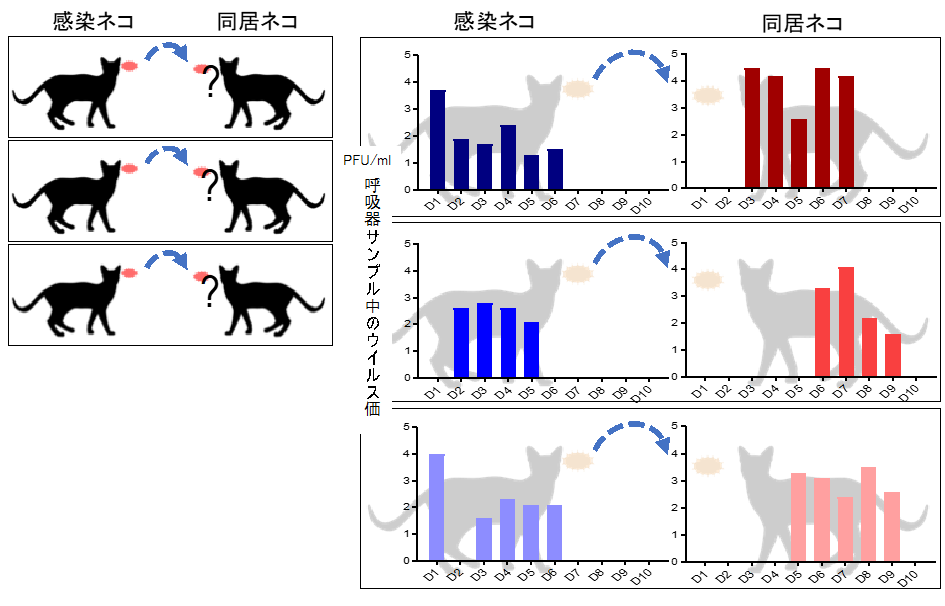
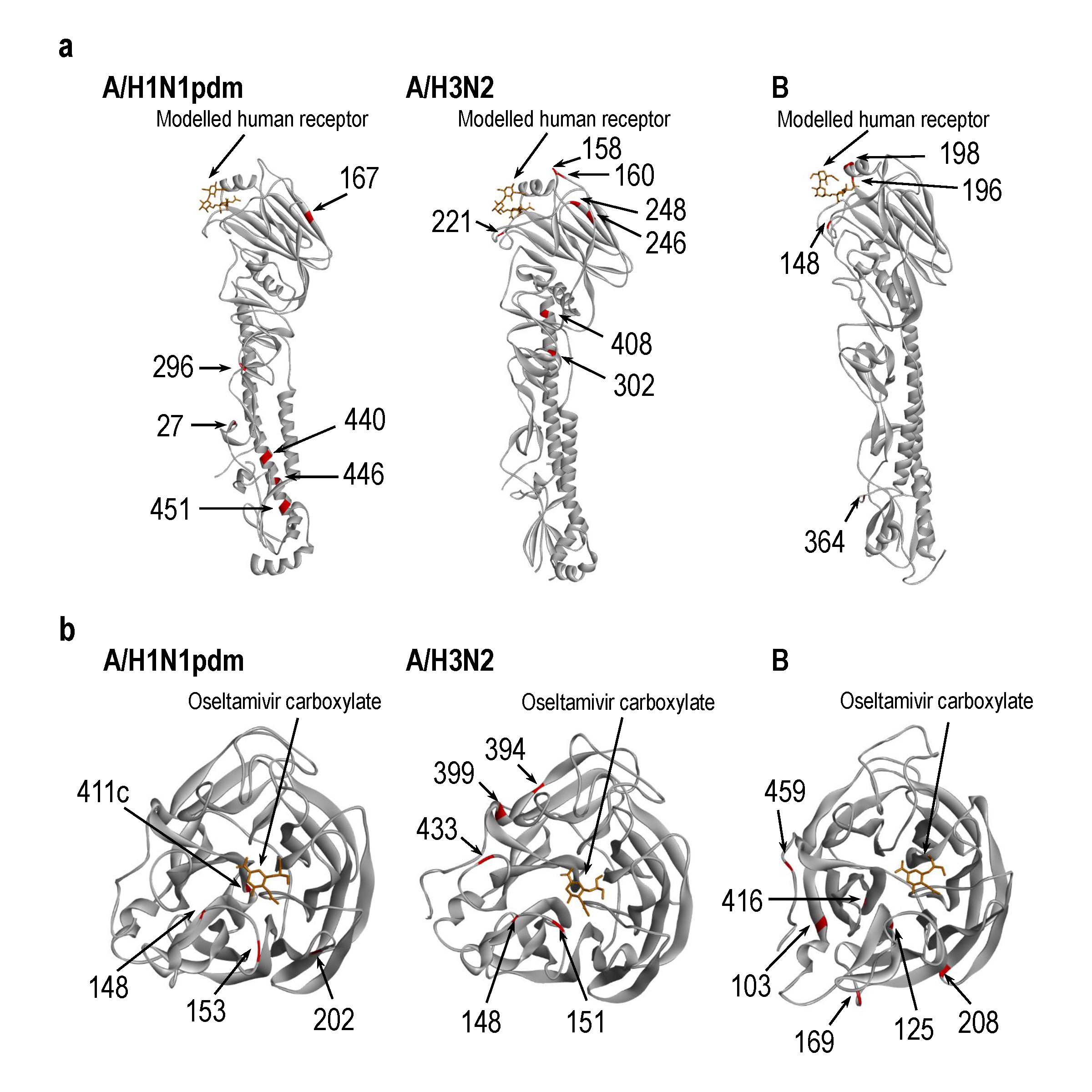
患者から分離されたゾフルーザ耐性インフルエンザウイルスの特性を解明
Influenza A variants with reduced susceptibility to baloxavir isolated from Japanese patients are fit and transmit through respiratory droplets
25 November 2019
 Localization of amino acid changes in HA and NA proteins
Localization of amino acid changes in HA and NA proteins
変異が入ることなく季節性インフルエンザウイルスを効率よく分離培養できる培養細胞株の開発に成功 ―細胞培養ワクチンへの応用―
A humanized MDCK cell line for the efficient isolation and propagation of human influenza viruses
29 April 2019
Subclade 2.2.1-Specific Monoclonal Antibodies That Recognize an Epitope in Antigenic Site A of Influenza A(H5) Virus HA Detected between 2015 and 2018.
2 April 2019
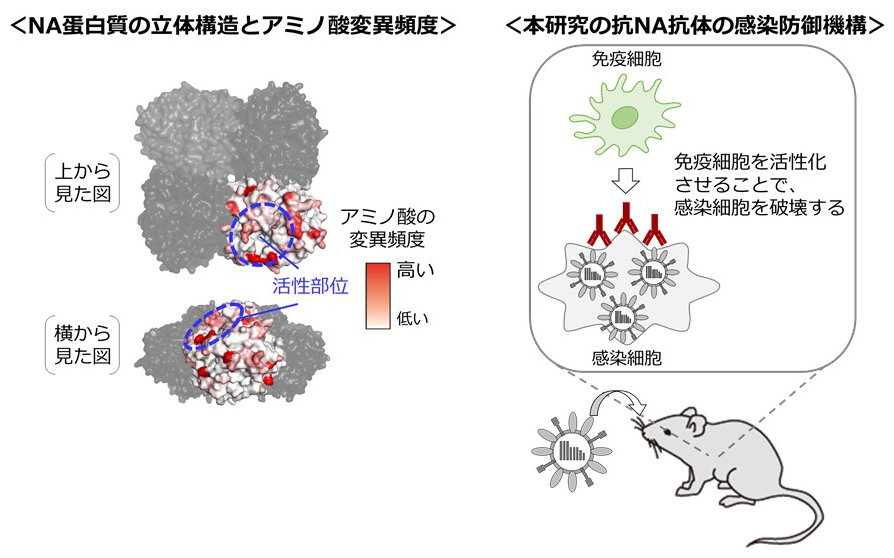
Antigenic drift originating from changes to the lateral surface of the neuraminidase head of influenza A virus.
18 March 2019
Highlights in 2018
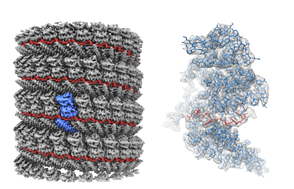 本研究で明らかになった核タンパク質-RNA複合体の構造
本研究で明らかになった核タンパク質-RNA複合体の構造エボラウイルス粒子のコア構造を解明
Cryo-EM structure of the Ebola virus nucleoprotein-RNA complex at 3.6Å resolution
17 October 2018
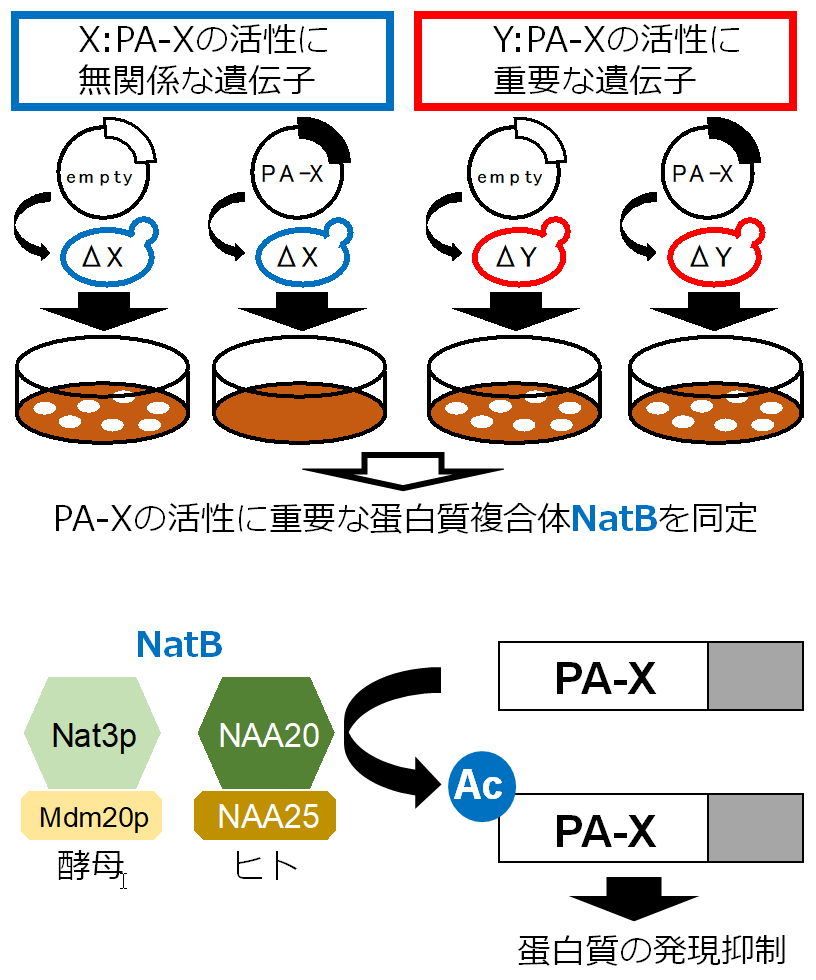
インフルエンザウイルスが宿主蛋白質の発現を抑制する仕組みを解明
N-terminal acetylation by NatB is required for the shutoff activity of influenza A virus PA-X
24 July 2018
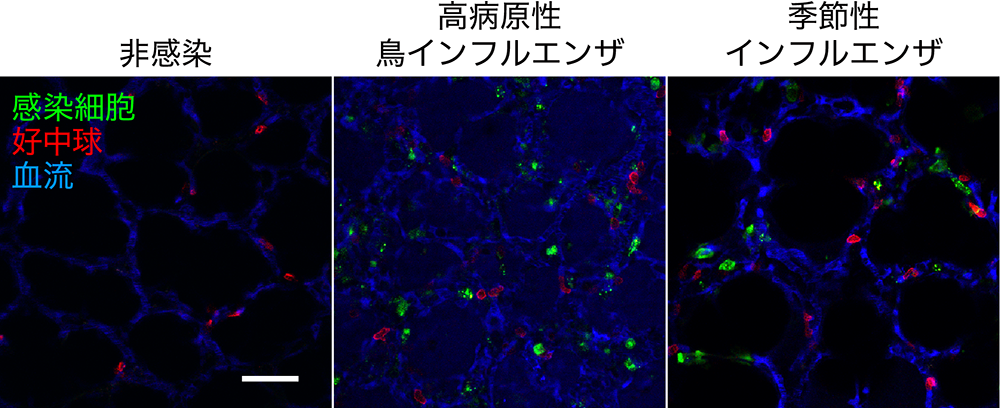
インフルエンザウイルスに感染した動物の体内を生きたまま観測
In vivo imaging of the pathophysiological changes and neutrophil dynamics in influenza virus-infected mouse lungs
25 June 2018
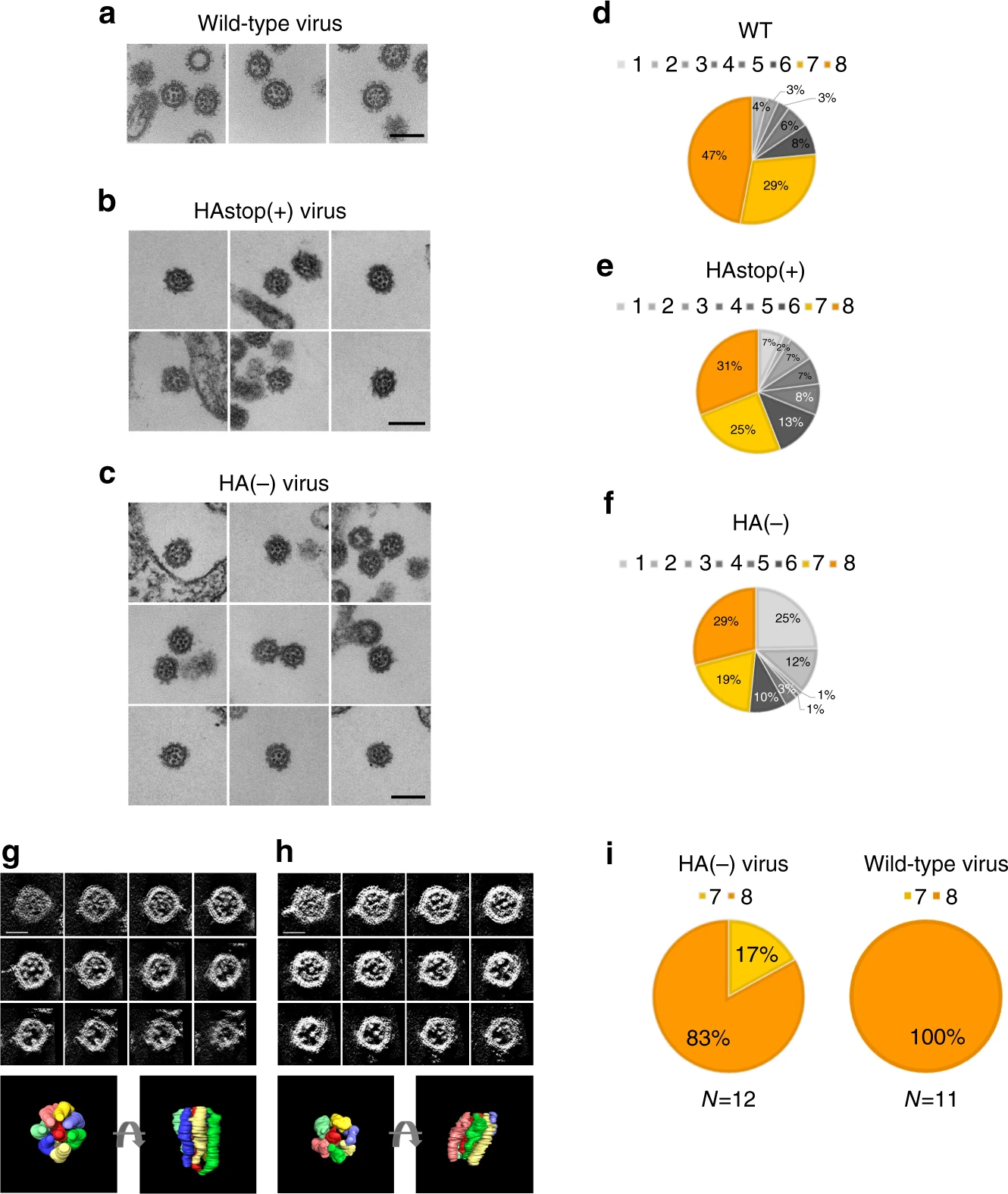 Electron micrographs of the HA(−), HAstop(+),
Electron micrographs of the HA(−), HAstop(+), and wild-type viruses. HA-MDCK cells were
infected with the wild-type virus, HAstop(+) virus,
or HA(−) virus, and ultrathin sections of budding
virions were observed by EM.
インフルエンザウイルスの遺伝の仕組みを解明
Importance of the 1+7 configuration of ribonucleoprotein complexes for influenza A virus genome packaging
4 January 2018
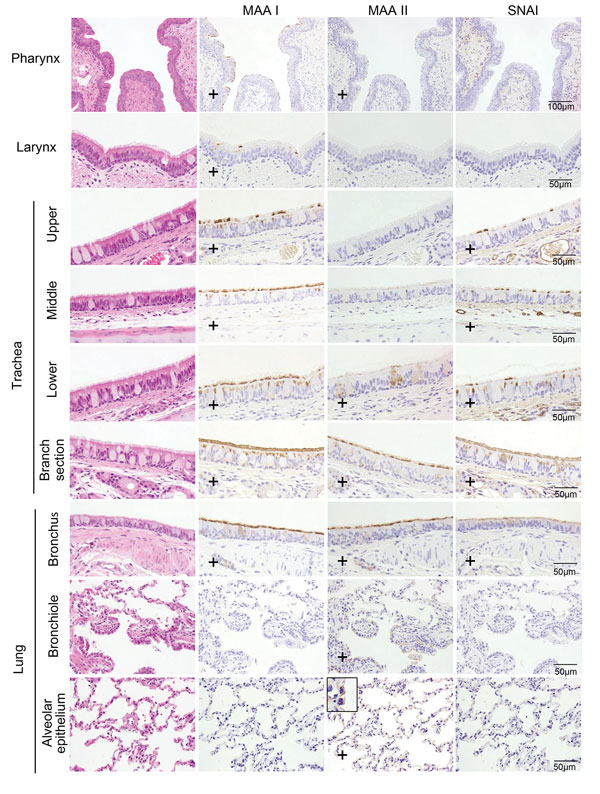 Distribution of α2,3- and α2,6-linked sialosides in the respiratory organs of a cat, New York, NY, USA.
Distribution of α2,3- and α2,6-linked sialosides in the respiratory organs of a cat, New York, NY, USA.
ニューヨークのネコで流行したH7N2インフルエンザウイルスの特性を解明
Characterization of a Feline Influenza A(H7N2) Virus
→ Emerging Infectious Diseases
January 2018
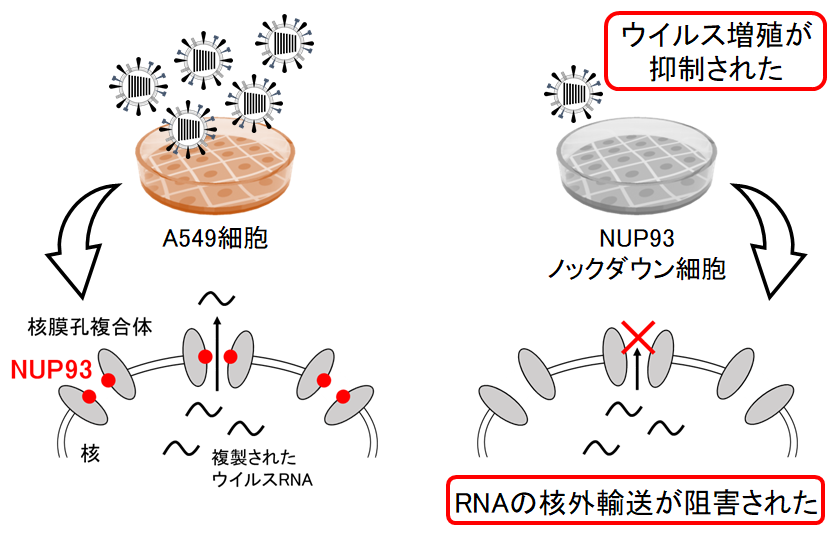
Host Factor Nucleoporin 93 Is Involved in the Nuclear Export of Influenza Virus RNA
24 July 2018

Development of an influenza rapid diagnostic kit specific for the H7 subtype
25 June 2018

The marmoset as an animal model of influenza: infection with A(H1N1)pdm09 and highly pathogenic A(H5N1) viruses via the conventional or tracheal spray route
9 May 2018

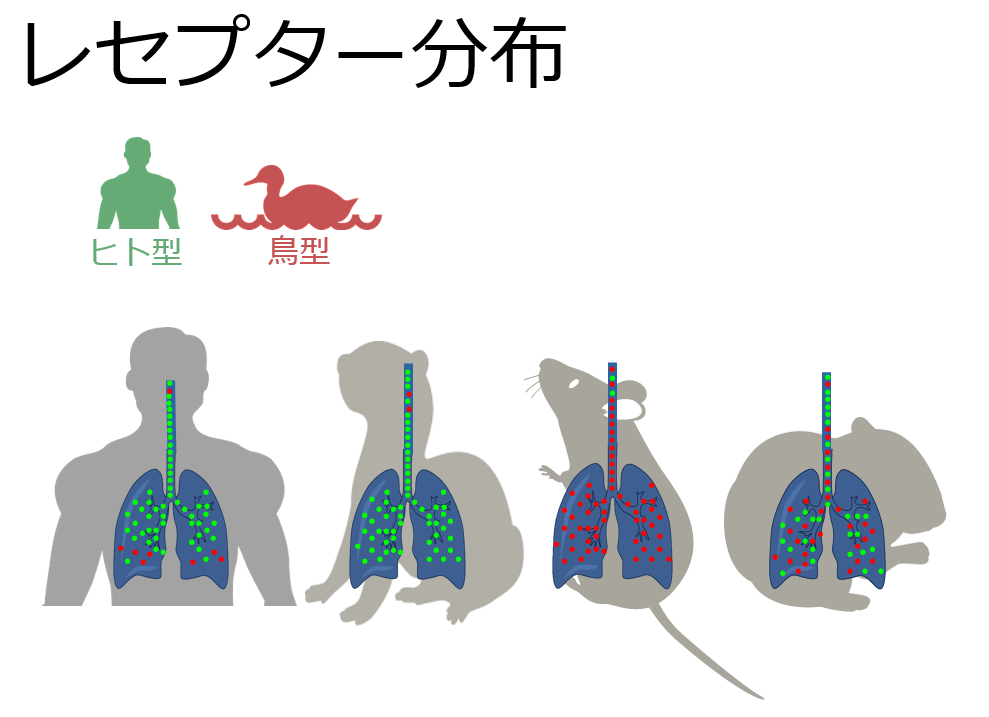
Enhanced Replication of Highly Pathogenic Influenza A(H7N9) Virus in Humans
→ Emerging Infectious Diseases
April 2018
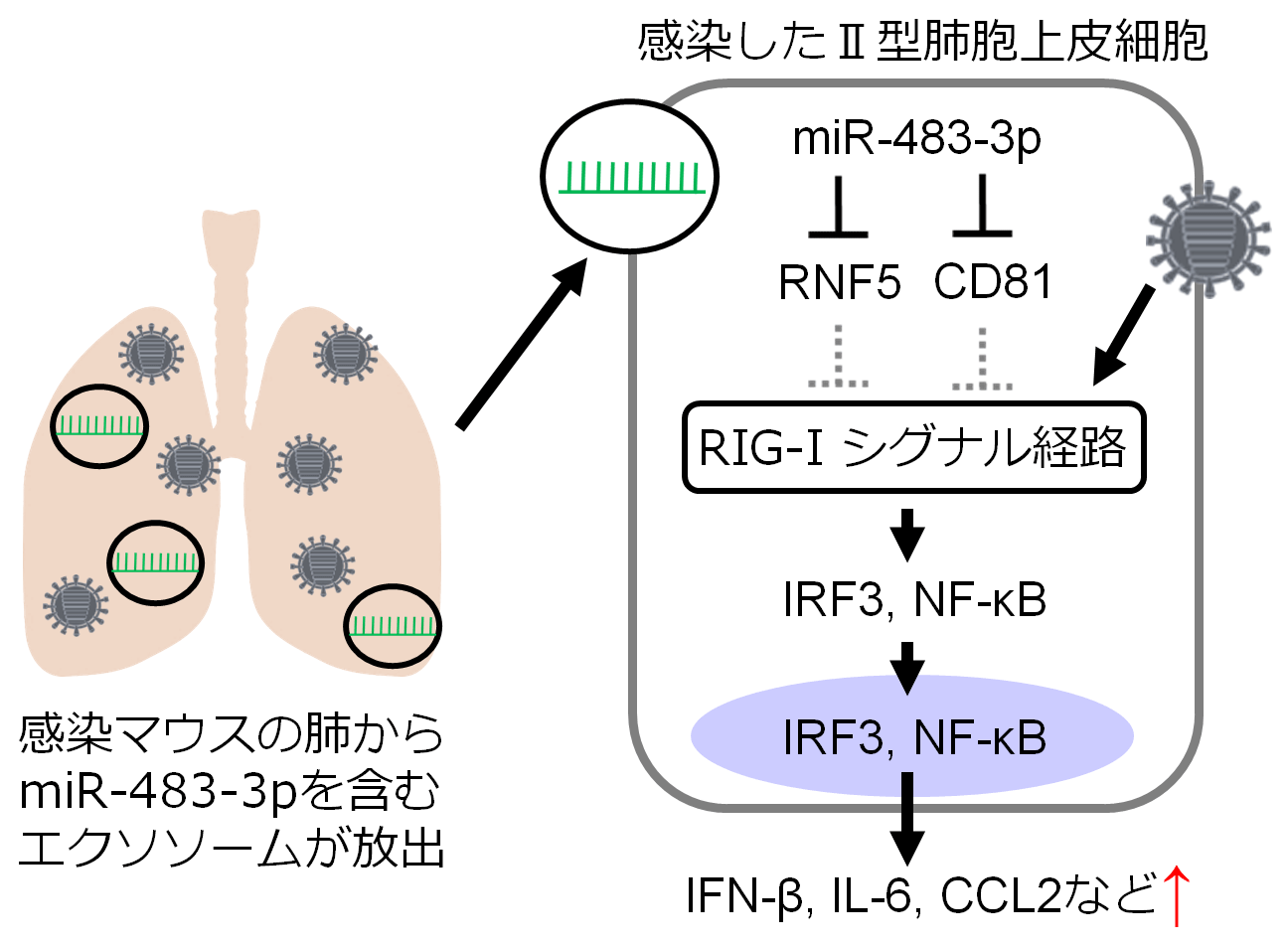
Lung-derived exosomal miR-483-3p regulates the innate immune response to influenza virus infection
→ The Journal of Infectious Diseases
24 January 2018

Identification of novel amino acid residues of influenza virus PA-X that are important for PA-X shutoff activity by using yeast
10 January 2018
Human protective monoclonal antibodies against the HA stem of group 2 HAs derived from an H3N2 virus-infected human
7 Feb 2018

Syrian hamster as an animal model for the study of human influenza virus infection
30 Jan 2018
Highlights in 2017 - 2012
Diversity of antigenic mutants of influenza A(H1N1)pdm09 virus escaped from human monoclonal antibodies
18 December 2017
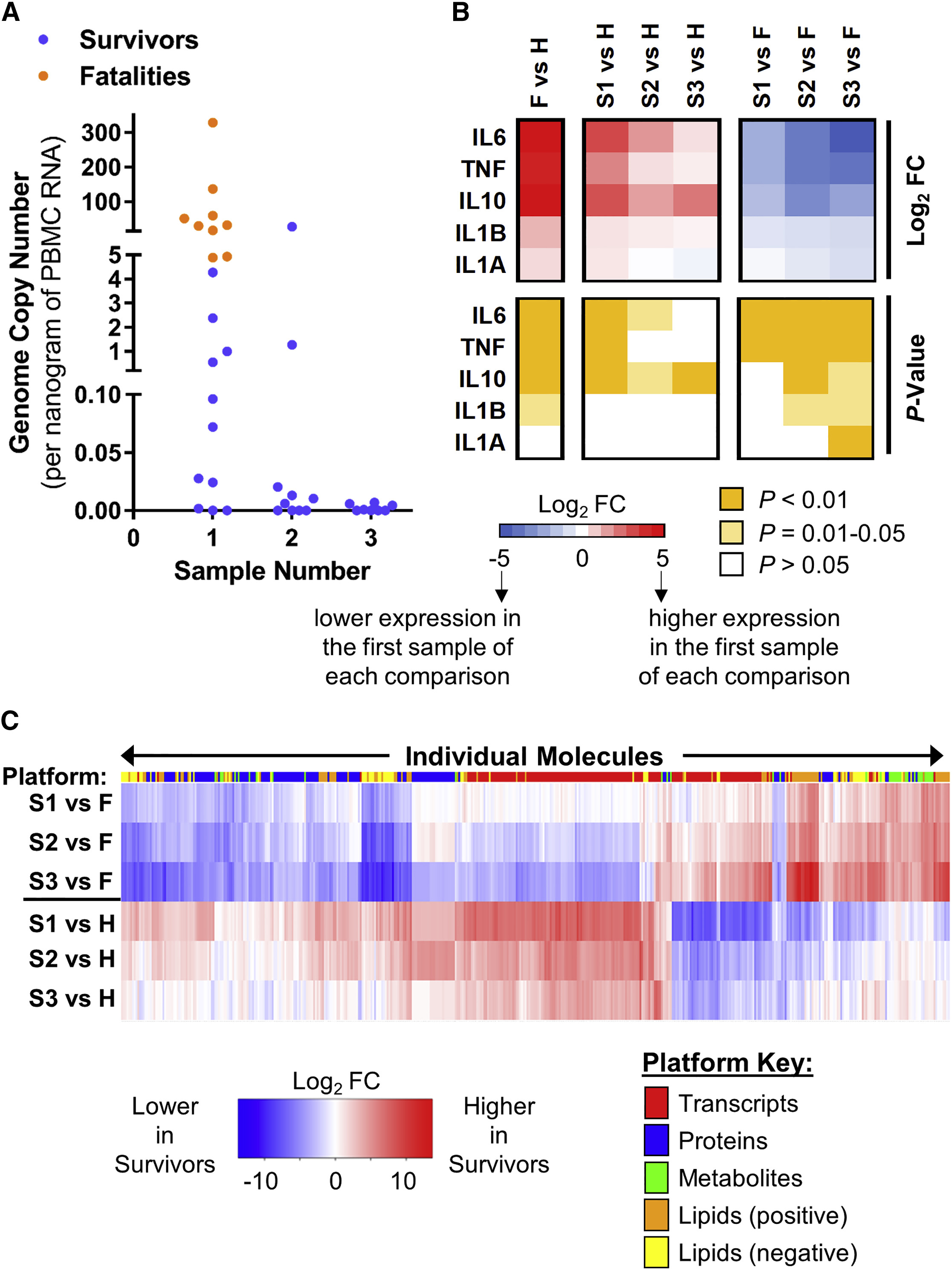 EVD Survivors and Fatalities Are Differentiated
EVD Survivors and Fatalities Are Differentiated by Viral Load and Host Responses
エボラ出血熱の重症化メカニズムの解明ならびに予後を予測するためのバイオマーカーを同定
Multi-Platform 'Omics Analysis of Human Ebola Virus Disease Pathogenesis
16 November 2017
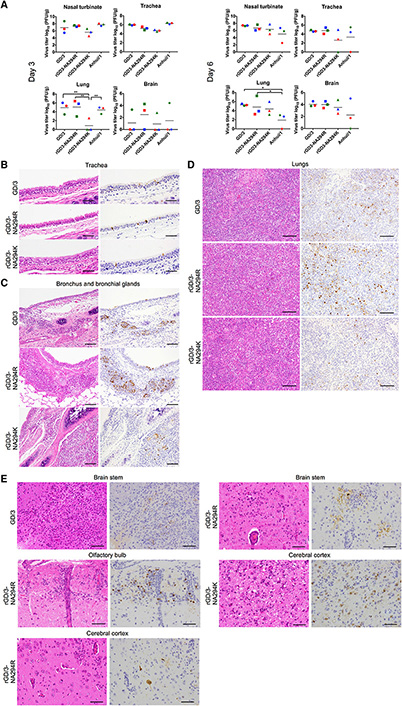 Virus Replication and Pathological Findings
Virus Replication and Pathological Findings in Infected Ferrets
中国の患者から分離された高病原性H7N9 鳥インフルエンザウイルスの特性を解明
A highly pathogenic avian H7N9 influenza virus isolated from a human is lethal in some ferrets infected via respiratory droplets
19 October 2017
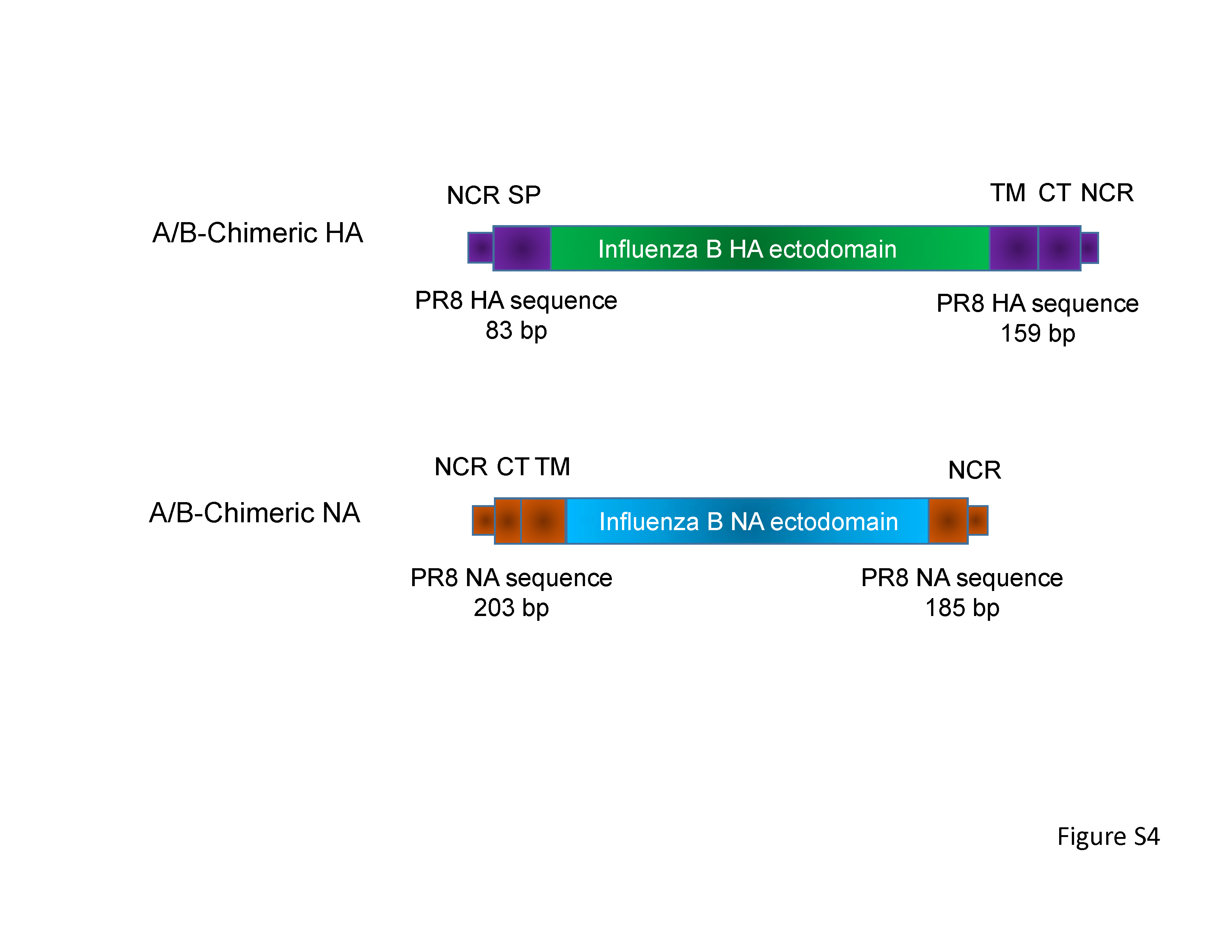 Schematic diagram of chimeric HA and NA genes
Schematic diagram of chimeric HA and NA genes
培養細胞で高い増殖能を持つB型インフルエンザウイルスの作出に成功 ~より迅速に、効率よく季節性ワクチンを製造することが可能に~
Development of high-yield influenza B virus vaccine viruses
5 December 2016
 Structural basis of antigenic changes
Structural basis of antigenic changes
季節性インフルエンザウイルスの抗原変異を予測する新規技術を開発 ~より有効なワクチン製造が可能に~
Selection of antigenically advanced variants of seasonal influenza viruses
23 May 2016
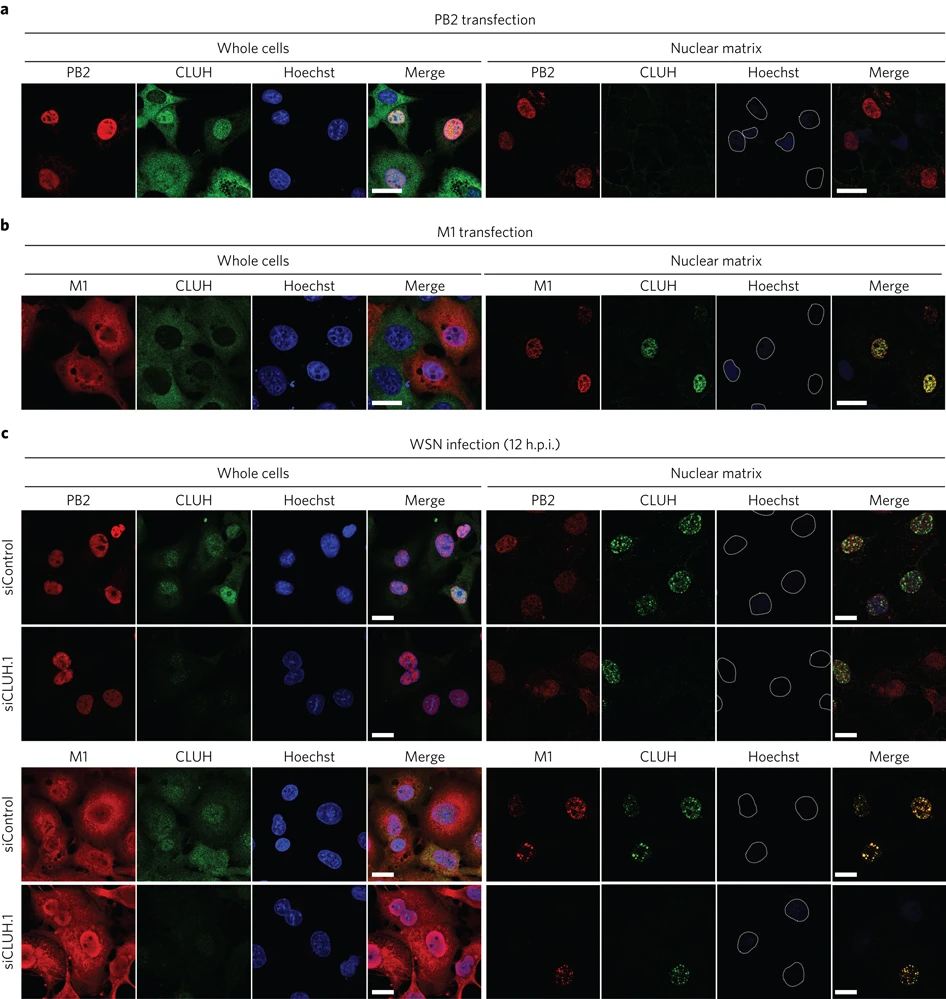
インフルエンザウイルスゲノムの核内動態に関わる宿主タンパク質を同定
The host protein CLUH participates in the subnuclear transport of influenza virus ribonucleoprotein complexes
16 May 2016
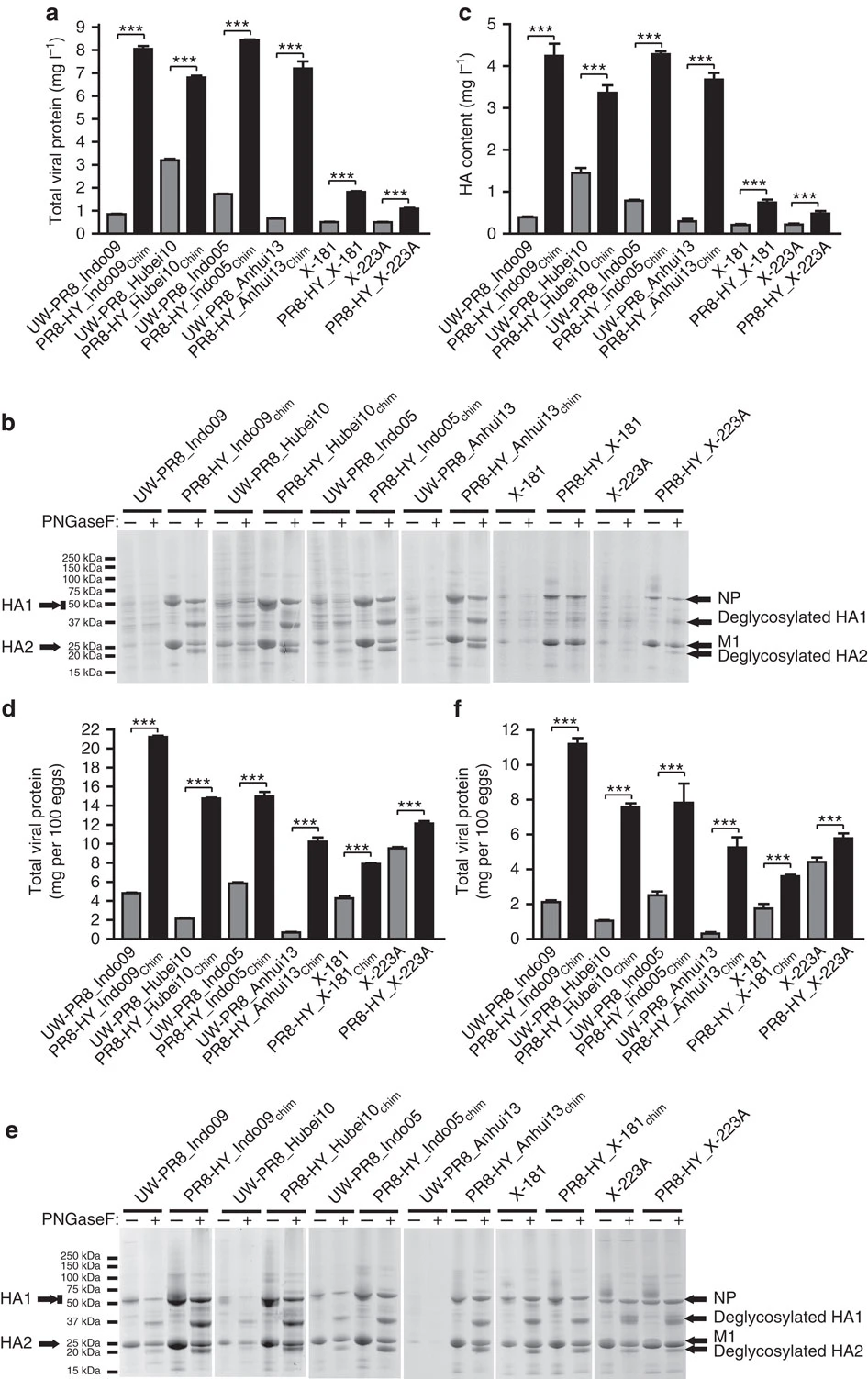 Evaluation of the total viral protein and HA
Evaluation of the total viral protein and HA content of PR8-HY candidate vaccine viruses
培養細胞での増殖能を大きく上昇させたインフルエンザウイルスの作製に成功 ~季節性ワクチン及びパンデミックワクチンの有効性上昇と迅速製造の道筋をつけた~
Development of high-yield influenza A virus vaccine viruses
02 September 2015
新しいエボラワクチンの開発に成功~ワクチンの有効性をサルで証明~
An Ebola whole-virus vaccine is protective in nonhuman primates
26 March 2015
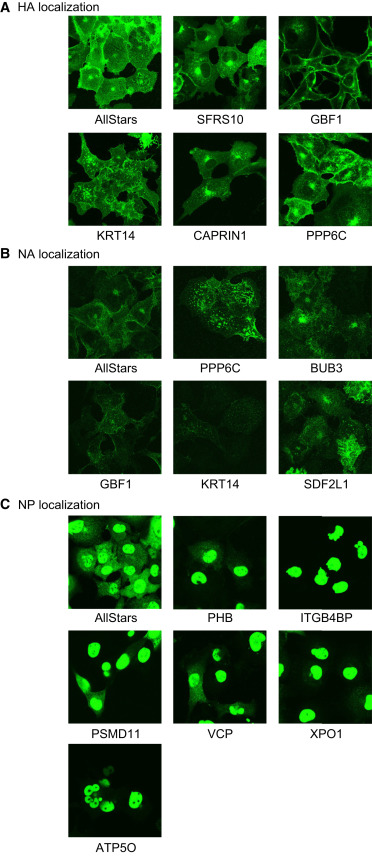 Effects of Selected siRNAs Targeting the 91
Effects of Selected siRNAs Targeting the 91 "Top Hits” on the Intracellular Localization
of Viral Proteins in Infected Cells
抗ウイルス薬開発へ向けたインフルエンザウイルスと宿主とのインタラクトーム解析
Influenza Virus-Host Interactome Screen as a Platform for Antiviral Drug Development
20 November 2014
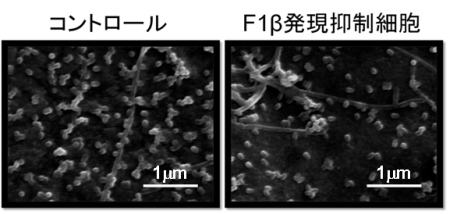 Scanning electron micrographs of
Scanning electron micrographs ofinfluenza virus-infected HEK 293 cells
インフルエンザウイルス粒子形成に関わる宿主因子の同定
F1Fo-ATPase, F-type proton-translocating ATPase, at the plasma membrane is critical for efficient influenza virus budding.
5 March 2012
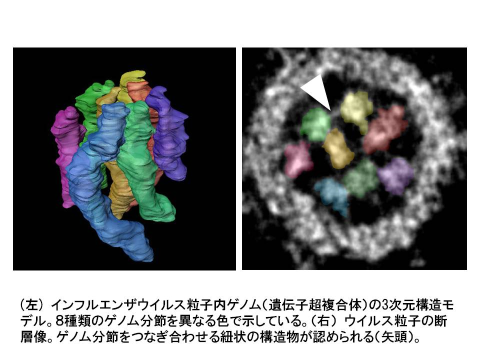
インフルエンザウイルス遺伝子の立体構造の解明
Three dimensional analysis of ribonucleoprotein complexes in influenza A virus
24 January 2012