研究活動
論文紹介
(Viruses 2019 Apr 2;11(4):E321.)
概要と感想
H5亜型の高病原性鳥インフルエンザウイルスは世界16カ国以上の国々の家禽や野鳥の間で流行しています。H5N1亜型のウイルスは稀にトリからヒトへ感染することが知られており、1997年以降860名に感染し、感染者の半数以上が死亡しています。H5亜型のウイルスは、膜蛋白質の一つであるヘマグルチニン (HA) の塩基配列の類似性に基づき、10個のCladeと多数のSubcladeに分類されています。近年は、Subclade 2.3.4.4に分類されるウイルスが東南アジア、ヨーロッパおよび北アメリカを中心に世界各国で、Subclade 2.3.2.1に分類されるウイルスは主にインドネシアで、Subclade 2.2.1に分類されるウイルスは主にエジプトで、Subclade 7.2に分類されるウイルスは中国で流行を続けています。
これまでに、H5亜型のHA蛋白質を認識するヒトモノクローナル抗体は22クローン報告されていました。これらは、HA蛋白質の抗原性のSubclade間での比較や、Subclade内での抗原性変化およびSubclade特異的に保存された抗原部位の解析に有用ですが、それらの詳細な解析にはより多種類のヒトモノクローナル抗体が必要です。
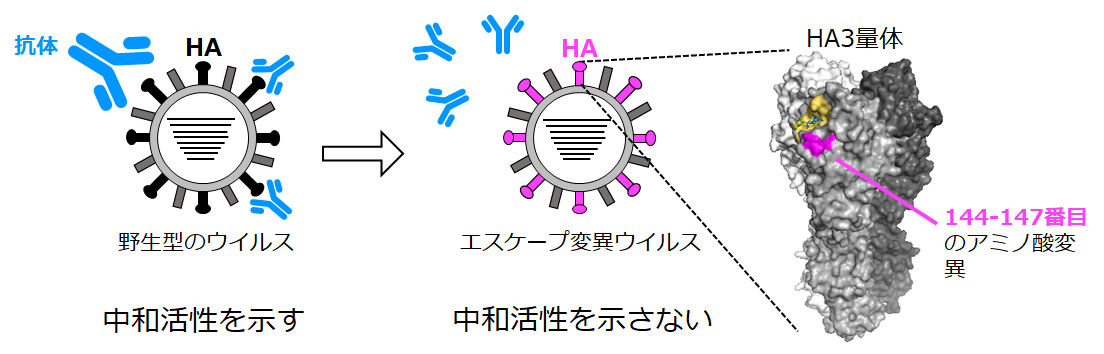
本研究では、Subclade 2.2.1またはSubclade 2.1.3.2に分類されるH5N1亜型のウイルスをワクチン抗原とするプレパンデミックワクチンを接種したボランティアの方々から、H5-HA蛋白質を特異的に認識するヒトモノクローナル抗体を15クローン得ました。そのうち9クローンが、Subclade 2.2.1に分類されるH5亜型のウイルスに対して赤血球凝集阻止 (HI)活性および中和活性を示しました。また、H5-HA蛋白質の144番目から147番目のアミノ酸変異または1アミノ酸の挿入が、抗体からのエスケープに重要なことが分かりました。HA蛋白質のAntigenic site Aに含まれるこの領域は、Subclade 2.2.1に分類されるウイルスに特異的な配列がSubclade内で高度に保存されていました。そのため、H5亜型ウイルスのSubclade間の抗原性の違いに関わる可能性が示唆されました。
研究を進めるにあたり、多数のハイブリドーマからヒトモノクローナル抗体を精製するまでの過程にはかなりの時間を要してしまいました。その分、実際に精製した抗体が、近年流行しているウイルス株に対して中和能をもち、主要なAntigenic siteを認識していることが分かったときは、努力が実を結んで嬉しかったです。初めてとなる論文投稿でしたが、修士2年間で取り組んだ内容を形にすることができて良かったです。
(Nature Microbiology 2019 Mar 18. doi: 10.1038/s41564-019-0401-1.)
概要と感想
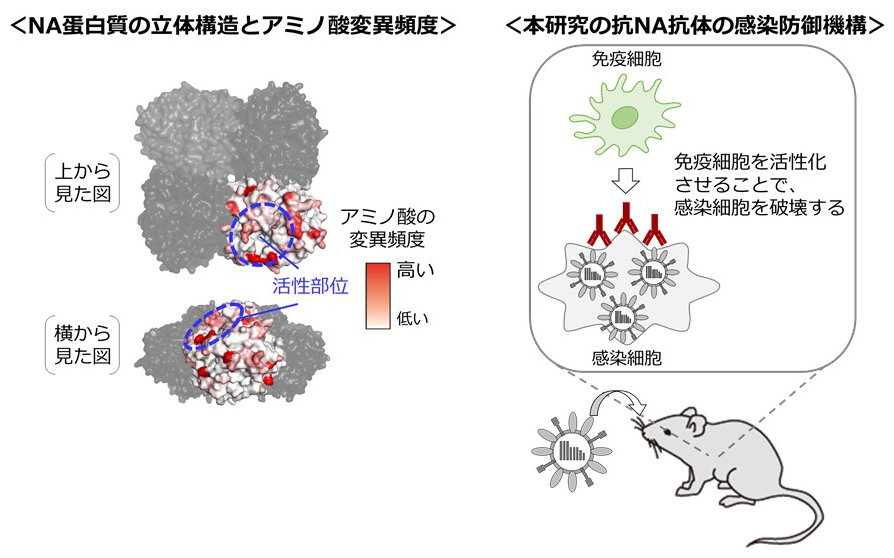
インフルエンザウイルスのノイラミニダーゼ(NA)蛋白質に対する抗体の機能解析は、NA蛋白質の酵素活性部位周辺を認識し、その酵素活性を阻害する活性(Neuraminidase inhibition; NI活性)をもつ抗体について行われてきました。NI活性をもつ抗体による感染阻害から逃れるために、NA蛋白質の活性部位周辺にはアミノ酸変異が生じています。しかしながら、活性部位から離れた「側面領域」にもアミノ酸変異が高度に蓄積された領域があります。この領域にアミノ酸変異が生じる理由は不明でした。
本研究では、NA蛋白質の側面領域を認識する抗体を解析することで、NI活性を持たない抗体がヒトの免疫細胞を活性化することで感染防御に寄与することを発見しました。さらに、NA側面領域のアミノ酸変異が、そのような抗体から逃れるために引き起こされていることを明らかにしました。現在NAの抗原性はNI活性をもつ抗体のみを対象として解析されていますが、本研究の成果はNA蛋白質の抗原性評価方法を改良する必要性を示唆しています。近年の研究により、NAに対する抗体が感染防御に重要であることが明らかとなっているため、本研究の成果はワクチン開発において重要な知見となります。
今回解析した抗体は、別の実験に必要な抗体をスクリーニングしている中で、たまたま見つかった抗体でした。そのため、当初は注目していませんでしたが、詳しく解析してみると大変興味深い結果が得られました。これこそ“セレンディピティ”と感じる最初の体験でした。
(Frontiers in Microbiology 2018 9:1675.)
概要と感想
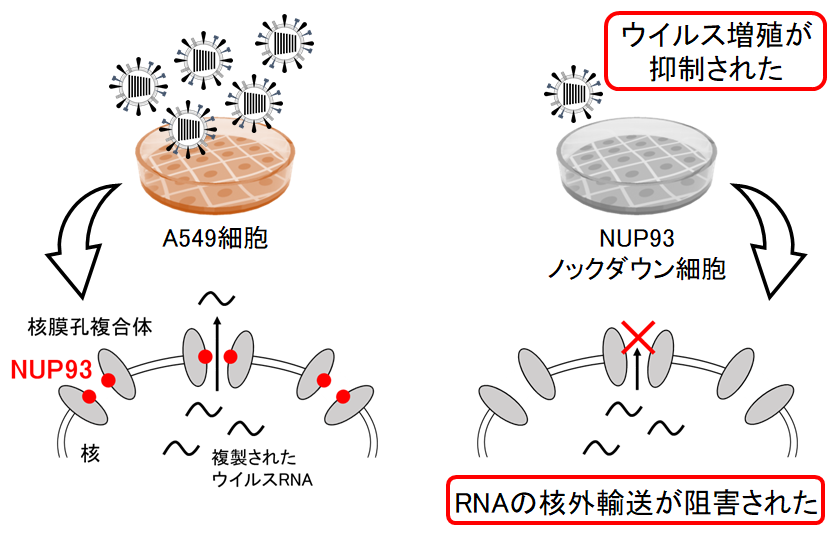
インフルエンザウイルスは宿主細胞の機能を利用して増殖します。当研究室では、網羅解析によって、ウイルスタンパク質と相互作用する多くの宿主因子を同定しました。本研究では、その中の1つであり、核膜孔複合体の構成タンパク質であるNUP93に着目しました。
siRNAを利用してNUP93をノックダウンした細胞にウイルスを感染させ、ウイルス増殖におけるNUP93の機能を解析しました。NUP93ノックダウン細胞では、ウイルスの増殖が抑制されました。さらに、NUP93ノックダウン細胞では、核内で合成されたウイルスRNAの核外輸送が阻害されることが示されました。以上の結果から、NUP93はインフルエンザウイルスのRNAの核外輸送において重要な機能を果たしていることが明らかになりました。
どの因子に着目して解析を進めるかは、RNAiスクリーニングで絞込んだのですが、多くの因子を対象にしたスクリーニングは根気がいる作業でした。ウイルスRNAの局在を調べるFISHの実験系の確立にも苦労しましたが、NUP93が関わるステップを特定することができて安堵しています。
(Frontiers in Microbiology 2018 9:1346.)
概要と感想
2013年の春以降、中国で低病原性H7N9鳥インフルエンザウイルスのヒトへの感染が報告されるようになりました。さらに、一部のH7N9ウイルスは高病原性になっています。H7N9ウイルスに感染したかどうかを迅速に判断し、患者さんを早期隔離することが、H7N9ウイルスの伝播を防ぎ、パンデミックを予防するための一つの手段となります。しかし、市販のインフルエンザ診断キットは、A型インフルエンザウイルスとB型インフルエンザウイルスを区別することができるものの、H7N9ウイルスと他のA型インフルエンザウイルスを区別することはできません。そこで、H7サブタイプを判別することが出来る迅速診断キットを開発しました。このH7キットは、H7N9ウイルスに感染した患者さんの早期発見に役立ちます。
H7N9ウイルスは今のところ日本では発生していません。そのため中国に行き、共同研究者にH7N9ウイルスに対する迅速キットの反応性を調べてもらいました。中国では皆とてもよく実験をしていて、活気と勢いを感じました。

(Frontiers in Microbiology 2018 9:844.)
概要と感想
インフルエンザの霊長類感染モデル動物には、カニクイザルやアカゲザルが用いられていますが、これらの動物は取扱いの煩雑さや飼育設備などの点から、実験に制限が生じます。そこで、新たな霊長類モデル動物として、マーモセットの可能性を検証しました。また、ウイルスの接種法も、従来法(経気管、経鼻、経眼、経口の同時接種)と気管噴霧法について比較検討しました。ウイルスを接種したマーモセットの肺をマイクロCTで観察した結果、従来法では肺の中心部に限局して強い炎症がおきていましたが、気管噴霧法では肺および気管全体に炎症が広がっていたことから、従来法より気管噴霧法が適していることが分かりました。気管噴霧法により高病原性鳥インフルエンザウイルスを投与した結果、体重減少、体温低下、食欲および活動量の低下が認められました。各種臓器のウイルス力価測定および病理解析では、呼吸器系組織のみならず、心臓、肝臓、腸管、脳など多数の臓器でウイルスの増殖が確認されました。これらの結果から、マーモセットはインフルエンザに対し高い感受性を示すことが明らかとなり、霊長類感染モデル動物になり得ると考えられました。
これまでにカニクイザルを用いた実験の経験はありましたが、マーモセットを扱うのは今回が初めてでした。マーモセットは、小さくてとても扱いやすく、小型のマイクロCTで生きたまま肺を観察することもでき、実験の幅が広がりそうだと感じました。

(Cell Reports 2018 Jul 24;24(4):851-860)
概要と感想
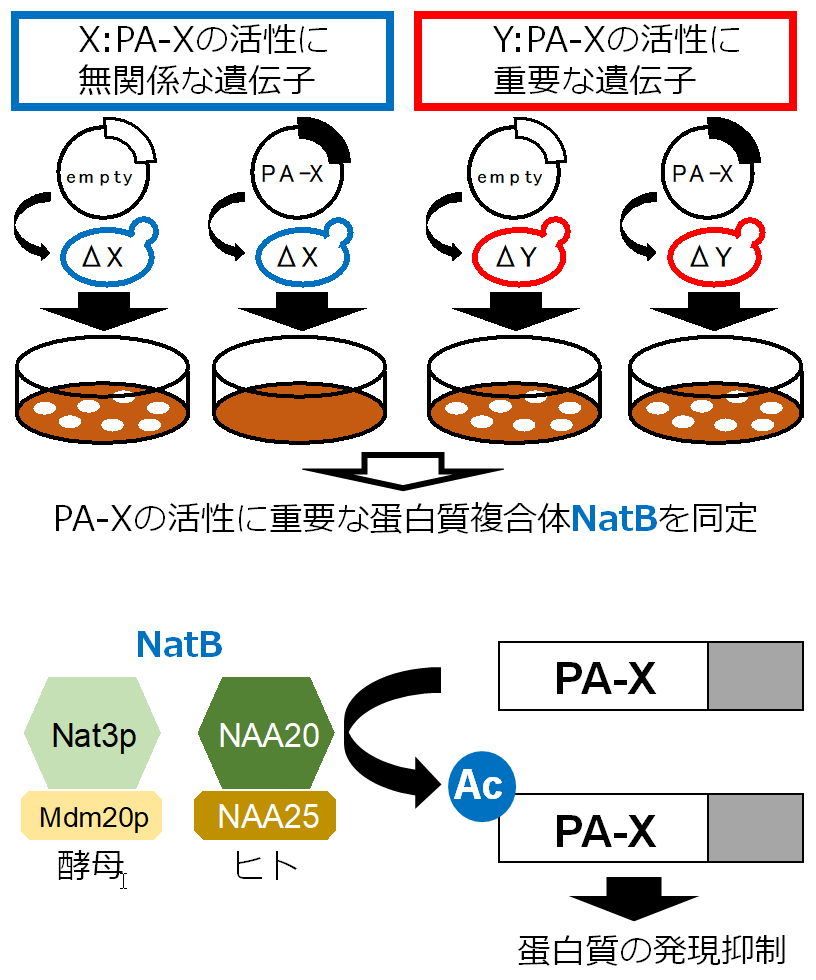
インフルエンザウイルスは感染細胞において、宿主蛋白質の発現を抑制することによりウイルスを排除しようとする防御反応を抑制します。この宿主蛋白質の発現抑制にインフルエンザウイルス蛋白質PA-Xが重要であることはわかっていましたが、PA-Xによる宿主蛋白質の発現抑制のメカニズムは明らかになっていませんでした。
本研究では、出芽酵母を用いたスクリーニングにより、PA-Xの活性に重要な宿主蛋白質としてNat3p (NAA20)とMdm20p (NAA25)を同定しました。これらの蛋白質はヘテロダイマーを形成し、蛋白質複合体NatBとして働くことが知られています。NatBは蛋白質の特定のN末端配列を認識し、そのN末端をアセチル化します。CRISPR/Cas9法により作製したヒトNatB欠損細胞や変異体PA-Xを用いた発現抑制活性の評価や質量分析を行った結果、NatBによるPA-XのN末端アセチル化がPA-Xの発現抑制活性に重要であることが明らかとなりました。
スクリーニングを行い候補遺伝子を同定した時はとてもわくわくしたのを覚えています。その後の解析にはかなり悪戦苦闘し、たくさん時間もかかってしまいましたが、なんとか成果をまとめることができ一安心しています。本研究で得られた知見がPA-Xの発現抑制機構のさらなる解析に役立つことを期待します。
(Proc Natl Acad Sci U S A. 2018 Jul 10;115(28):E6622-E6629)
概要と著者感想
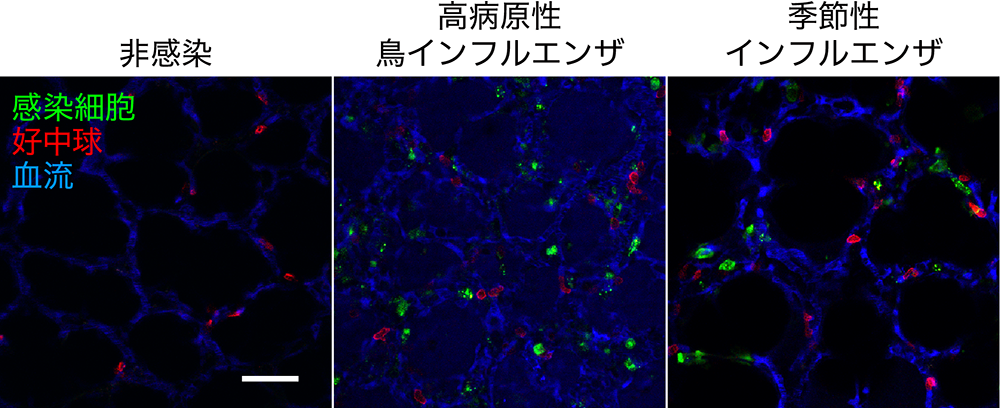
インフルエンザは、時として致死性の肺組織障害を引き起こすため、医学・獣医学・公衆衛生上の対策が必須な呼吸器感染症です。インフルエンザウイルスに感染した肺では、免疫系の活性化をはじめ様々な宿主応答が誘導されると考えられていますが、従来の固定標本などを用いた解析では、細胞の動きや血液の流れなどの時間軸を持った情報を得ることはできませんでした。
本研究では、2光子励起顕微鏡を用いた生体イメージングシステムを構築することで、インフルエンザウイルスに感染したマウスの肺における免疫細胞の動きや血液の流れをタイムラプス像として撮影することに成功し、血流速度、血管透過性の変化、免疫細胞の移動速度などの観測、および、新たな病態生理学的なパラメーターとしての定量化解析を行うことができました。さらに、このイメージングシステムをバイオセーフティーレベル3 (BSL3) の施設に設置することで、季節性ヒトインフルエンザウイルス(H1N1)のみならず高病原性鳥インフルエンザウイルス(H5N1)に感染した動物の観察が可能となり、病原性の異なるウイルス株を比較解析することができました。
本研究で確立したインフルエンザウイルス感染肺の生体イメージングシステムは、他の肺疾患の解析にも応用が可能であり、様々な呼吸器疾患の病態解明にも役立つことが期待されます。
2光子励起顕微鏡を用いた生体イメージング法は、ウイルス学以外にも免疫学や外科学、顕微鏡の操作法や画像解析手法など広範囲の知識や技術が必要でしたので、ひとつひとつトラブルシューティングしていくのが大変でしたがその分やりがいもありました。インフルエンザウイルスに感染した肺を初めて撮影できた時は感動的でした。
(Emerg Infect Dis 2018 Apr;24(4):746-750.)
概要と著者感想
2013年3月、中国においてH7N9亜型の鳥インフルエンザウイルス(H7N9ウイルス)の鳥から人への最初の感染が報告された。2013年以降、感染者が毎年報告されており、2017年5月までに感染者1486名、死者571名に達している。H7N9ウイルスは人から人への飛沫核感染が可能なものの、その効率は高くない。2016-2017年シーズンには、ヒトおよび鶏においてHAの開裂部位に複数の塩基性アミノ酸を持つ高病原性H7N9ウイルスが確認され、公衆衛生上の大きな懸念となっている。
一般に、鳥類から分離された鳥インフルエンザウイルスは人の体内では増殖しがたい。ウイルスポリメラーゼPB2のアミノ酸変異(Q591K、E627KまたはD701N)により、鳥インフルエンザウイルスは哺乳類で増殖しやすくなる。人から分離された多くのH7N9ウイルスには上記の変異がみられるが、これらを持たない人分離株が近年報告されており、そのようなウイルスがどのようなアミノ酸変異をポリメラーゼ蛋白質に獲得し、哺乳類で効率よく増殖するようになったのかを調べた。
2013年に分離されたH7N9ウイルスA/Anhui/1/2013株と2016年に分離されたPB2に既知の適応変異を持たないH7N9ウイルスA/Guangdong/17SF003/2016(GD/003)株を比較することで、ウイルスの効率良い増殖に関わるアミノ酸変異の同定を試みた。GD/003株の哺乳動物での高いポリメラーゼ活性、効率良い増殖および高い病原性には、PB2の482番目のアルギニン(R)、588番目のバリン(V)およびPAの497番目のアルギニン(R)がそれぞれ重要であることが分かった。今後、これらのアミノ酸を持ったウイルスの拡大に注意が必要である。
高病原性H7N9ウイルスの出現が分かってから全ての実験が完了するまで、6カ月という短期間で解析を完了することが出来た。実験計画の重要性を改めて知ることが出来た。

(The Journal of Infectious Diseases. 2018 Apr 11;217(9):1372-1382.)
概要と著者感想
エクソソームは細胞外小胞のひとつで、細胞間を移行して情報伝達を行います。近年の研究で、宿主がウイルスに感染した時に、感染細胞や周囲の免疫細胞から放出されたエクソソーム内の蛋白質や機能性RNAを介して、抗ウイルス応答が生じることが報告されています。これまでインフルエンザウイルスの感染時に放出されるエクソソームの機能は分かっていませんでした。
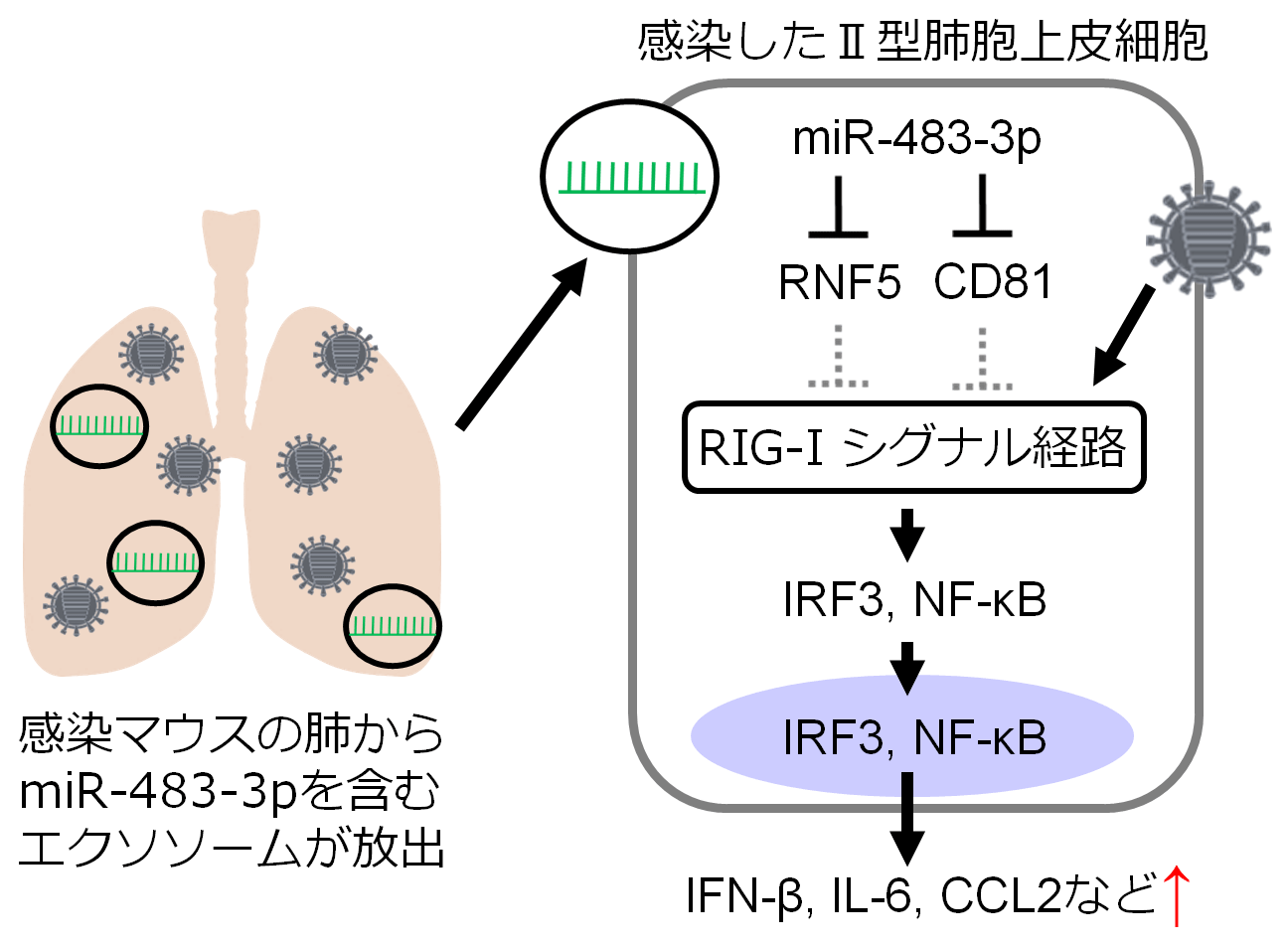
本研究では、インフルエンザウイルスを感染させたマウスの気管支肺胞洗浄液(BALF)中のエクソソームの解析を行い、肺から放出されたエクソソーム内に含まれる様々な種類のmicroRNAを同定しました。これらのmicroRNAの機能を明らかにするために、マウスのⅡ型肺胞上皮細胞を用いて解析した結果、いくつかのmicroRNAは抗ウイルス応答に重要な分子であるIFN-βや炎症性サイトカインの発現を増強することがわかりました。最も高い発現増強作用を示したmiR-483-3pは、RIG-Iシグナル経路の負の制御因子として知られるRNF5や新規の制御因子として同定したCD81の発現を抑制し、転写因子であるIRF3やNF-κBの核移行を促進することで、IFN-βや炎症性サイトカインの発現を増強していることが明らかになりました。本研究の結果から、インフルエンザウイルス感染時には感染局所においてエクソソーム内microRNAを介した宿主応答が生じている可能性が示唆されました。
今回同定したmicroRNAがインフルエンザウイルス感染に対してどのような機能を持っているのかは不明でした。マイクロアレイデータからのmicroRNAの選抜、スクリーニング解析に使用する細胞の検討、どのような機能に注目するか、など各ステップで時間がかかりました。miR-483-3pの機能が少しずつ明らかになっていくときには、時間をかけて選んできたものが実を結んだような思いで、嬉しかったのを覚えています。
(Scientific Reports 7, Article number: 17735 (2017), doi:10.1038/s41598-017-17986-8)
概要と著者感想
2013年以降のA(H1N1)pdm09亜型インフルエンザウイルスでは、ウイルス膜蛋白質ヘマグルチニン(HA)の166番目のアミノ酸が変異することで、ヒト抗体の中和から逃れていることが報告されています。しかしながら、HAの166番目のアミノ酸周辺を認識するヒト抗体から逃れることに、166番目以外のアミノ酸変異が寄与するかは不明でした。本研究では、HAの166番目のアミノ酸周辺を認識するヒトモノクローナル抗体とともにウイルスを継代することで抗原変異株を人工的に作製しました。その結果、HAの166番目の他に129、165番目のアミノ酸変異によって、ウイルスは抗体の中和を逃れることがわかりました。さらに、抗原変異株は親株と同程度以上の増殖性を示すことから、129および165番目のアミノ酸に変異を有する抗原変異株も、166番目のアミノ酸変異株と同様に流行株となる可能性が示唆されました。本研究は、抗体を用いた抗原変異株の解析が抗原変異の予測に貢献できることを示しています。
このような実験を行っているとウイルスは様々な変異を獲得することが確認できますが、自然界では166番目のアミノ酸変異といった特定の変異を持つウイルスのみが流行します。自然界では、抗体の中和から逃れることや増殖性の他にも様々な要因が重なり合ってウイルスの進化を決めており、それを再現することは大変奥が深く、興味深いテーマであると感じました。
(Virology 2018 Jan 10; 516: 71-75)
概要と著者感想
インフルエンザウイルス蛋白質PA-Xは宿主およびウイルス蛋白質の発現を抑制するshutoff活性をもちます。
酵母にPA-X発現プラスミドを導入しPA-Xを発現させようとすると、PA-Xのshutoff活性が酵母の生育を阻害するため、酵母はほとんどコロニーを形成しません。しかし稀に、PA-X発現プラスミドを導入しても酵母がコロニーを形成することがあります。形成されたコロニーを解析すると、PA-Xのshutoff活性が低下するような変異が導入されていることがわかりました。本研究では、この現象を利用することで、PA-Xのshutoff活性に重要なアミノ酸を新たに22残基同定しました。
ウイルスの研究にも関わらずウイルスを使わず、酵母を多く用いている一見風変わりな研究なのですが、酵母は実験ツールとしてとても有用なのでこれからもウイルス研究に応用していきたいと思います。

(J Infect. 2018 Feb;76(2):177-185.)
概要と著者感想
A型インフルエンザウイルスの糖蛋白質HAは、抗原性の違いから2グループ18亜型に分類される。HAは構造上headとstemの2つの領域に分けられ、head領域の抗原性は亜型間で大きく異なるのに対し、stem領域の抗原性はグループ内で高度に保存されている。これまでにHAのstem領域に対する抗体が、様々な亜型のインフルエンザウイルスに対して感染防御効果を示すことから、ノイラミニダーゼ阻害薬やポリメラーゼ阻害薬と共に、新型インフルエンザウイルスによるパンデミック時の治療薬として期待されている。
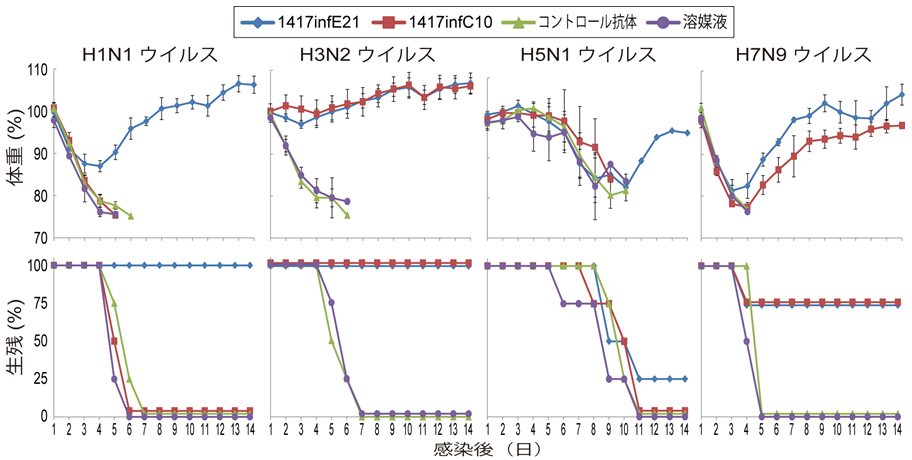
我々がH3N2ウイルス感染患者より、H3-HAやH7-HAが分類されるグループ2のHA Stem領域を認識する2種類のヒトモノクローナル抗体(1417infE21および1417infC10)を樹立した。この2クローンの抗体は、H3N2ウイルスおよびH7N9ウイルスの致死的感染からマウスを防御することが示された(図)。
我々が以前に報告した抗体(S9-1-10/5-1)と今回の抗体を組み合わせることで、より有効な抗体治療法となることが期待される。
実験とその結果に関しては想定通りであったものの、他の論文との差別化に苦労した。
(J Virol. 2018 Jan 30;92(4))
概要と著者感想
インフルエンザウイルスの感染実験モデル動物には、フェレットとマウスが広く用いられています。しかし、フェレットは取扱いの煩雑さや飼育設備、費用などの点から、最適なモデル動物とは言い難く、マウスはヒトから分離されたウイルスに対する感受性が低いため、ウイルスをマウスで継代してマウス馴化株を作らなければなりません。特に近年ヒトから分離されるH3N2亜型のインフルエンザウイルスは、マウスに感染させても増えにくく、マウス馴化株の作製すら困難です。そこで、フェレットおよびマウス代わるモデル動物として、シリアンハムスターの可能性を検証しました。その結果、ヒトから分離したウイルスは馴化しなくてもハムスターで増殖できることが分かりました。さらに飛沫伝播について調べたところ、B型は飛沫伝播しませんでしたが、H1N1亜型のウイルスはハムスター間でよく飛沫伝播し、H3N2亜型も2株中1株が飛沫伝播しました。これらの結果から、シリアンハムスターはインフルエンザの感染実験モデル動物になり得ると考えられました。ハムスターは、搬入直後は緊張しているのか若干攻撃的ですが、お世話をしているうちにどんどん慣れてきます。慣れてくると仰向けになって寝たりするので、死んでしまったのかと驚かされたことが何度もあります。扱いやすい動物なので、広く使われると良いと思います。

(J Infect Dis. 2017 Nov 25. doi: 10.1093/infdis/jix606)
概要と著者感想
免疫不全患者は、インフルエンザウイルスに感染した場合、抗ウイルス薬による治療にもかかわらずウイルスが体内に長期間持続し、予後は不良、かつ耐性ウイルスが検出される事例が数多く報告されています。また、ヌードマウスは先天的に無毛で、胸腺を欠くため免疫系が阻害されており、免疫不全モデルとして使用されています。本研究では、ヌードマウスを用い、 NA阻害剤、ポリメラーゼ阻害剤および両剤併用の効果について検討しました。
致死量のインフルエンザウイルスをヌードマウスに接種した場合、薬剤を長期間併用投与することにより多くのマウスは生残しましたが、投与を中止すると徐々に効果が消失し、最終的にはすべてのマウスが死亡しました。また、マウス肺からはNA阻害剤耐性ウイルスが検出されました。このことは、免疫不全患者にとっては、さらに効果的な治療が必要であることを示しています。
ヌードマウスは非常に面白い結果を出してくれるので、今後も引き続き実験を継続していきます。
(EBioMedicine. 2017 Mar;17:182-191.)
概要と著者感想
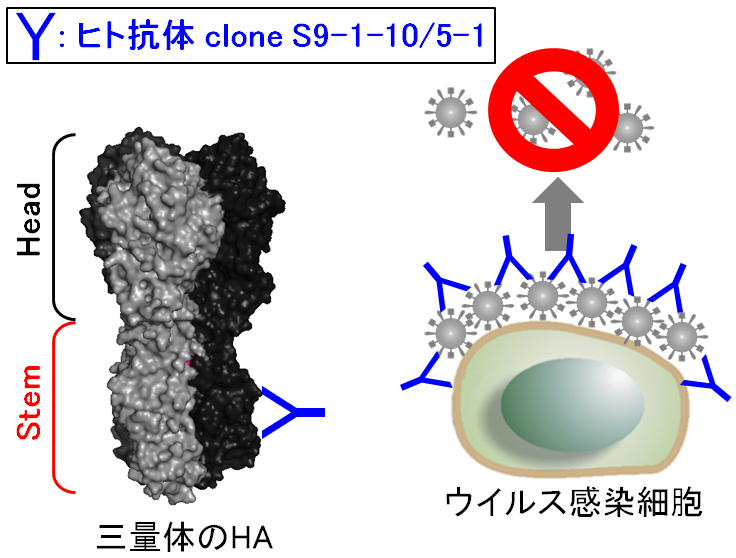
A型インフルエンザウイルスのHAは抗原性の違いから2グループ18亜型に分類される。HAは構造上headとstemの2つの領域に分けられ(図の左)、head領域の抗原性は亜型間で大きく異なるのに対し、stem領域の抗原性はグループ内で高度に保存されている。stem領域には2つのグループに共通な抗原部位も存在する。
我々がH5N1プレパンデミックワクチン接種者から樹立したヒトモノクローナル抗体S9-1-10/5-1は、全ての亜型のHAを認識し、主にウイルス粒子の放出過程を阻害していることを見出した。S9-1-10/5-1は、現在ヒトでの感染が確認されているH1N1pdm09、H3N2、H5N1およびH7N9ウイルスの致死的感染からマウスを防御することを確認した。
以上の結果から、S9-1-10/5-1がこれまで報告された抗stem抗体とは異なるメカニズムでウイルスの増殖を阻害していることを明らかにすると共に、この抗体を利用した治療が有用である可能性を示すことに成功した。
河岡研におけるヒト抗体を用いた研究の最初の論文であったため、全てが初めてのことで手探りで進めていた。当初考えていたよりも大変ではあったが、最終的な結論が想定と違う形に行きついた。
(mBio 2016 7(5):e01248-16.)
概要
A型およびB型インフルエンザウイルスは、8本に分節化したRNAをゲノムとして持っています。それぞれのRNA分節は核タンパク質およびRNA依存的RNAポリメラーゼと結合し、ウイルス粒子内でRNPと呼ばれる棒状の構造物を形成しています。感染細胞から出芽する際、子孫ウイルスが感染性を持つためには、ウイルス粒子内に8種類すべてのRNPを取り込む必要があります。しかし、8種類8本のRNPがどのようにウイルス粒子内に取り込まれているのか、詳細な機構はわかっていません。
これまでに、A型実験室株では、ウイルス粒子内に8本のRNPが選択的に取り込まれることが示されています。しかし、臨床分離株やB型ウイルス粒子内にRNPがどのように取り込まれるかは明らかになっていません。
本研究では、透過型電子顕微鏡を用いて電子線トモグラフィーを行うことで、A型およびB型ウイルスの実験室株と臨床分離株のウイルス粒子内に取り込まれたRNPの配置と本数を解析しました。
その結果、A型およびB型ウイルスは、株に依らず、一部のウイルス粒子が7本以下のRNPを取り込むものの、ほとんどのウイルス粒子が8本のRNPを取り込むことが明らかになりました。
本研究で得られた知見は、株や型に依らないインフルエンザウイルスの増殖メカニズムを紐解く重要な手がかりとなることが期待されます。
インフルエンザウイルスを様々な角度から電子顕微鏡で撮影し(A)、得られた像をコンピューターで処理することで(B)、一つのインフルエンザウイルス粒子全体を再構築することができる(C)。この手法を用いることで、ウイルス粒子内に取り込まれたRNPの配置と本数を知ることができる。
(Nat Microbiol. 2016 pii: 16062.)
概要
宿主細胞の核内で複製されたインフルエンザウイルスゲノムが、核内で輸送されるのに関わる宿主のタンパク質としてCLUH(clustered mitochondria protein homolog)を同定しました。
ウイルスゲノムの複製とウイルスゲノムの核外輸送複合体の形成は、核内で起きると報告されてきました。ところが、ウイルスの増殖に必要な「複製」と「複合体形成」が核内の同じ場所で起きているかどうかは不明でした。本研究において、新規に複製されたウイルスゲノムは、核内を移動した後に、核外輸送複合体が形成される領域へ到達することが明らかとなりました。
CLUHは通常細胞質に存在しますが、ウイルス感染により核内へと移動し、複製されたウイルスゲノムを運ぶ役割に関わることがわかりました。本研究成果は、ウイルス感染細胞の核内でのみ見られる現象を明らかにしたものであり、特異的なインフルエンザ治療薬開発のターゲットになることが期待されます。

(J Virol. 2015 Jun 3. pii: JVI.01132-15.)
概要
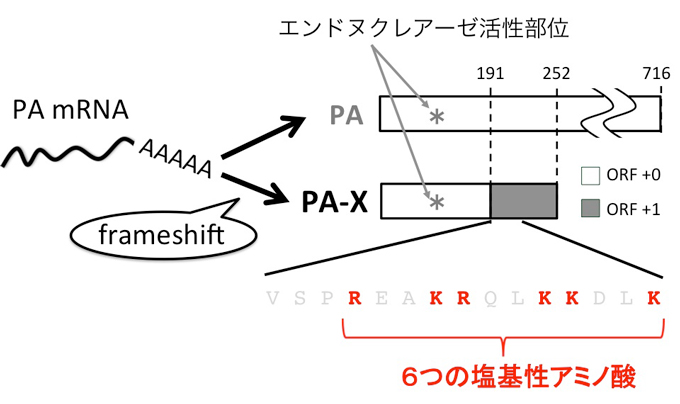
新規インフルエンザウイルス蛋白質PA-Xは、PAをコードするmRNAからPA蛋白質が翻訳される過程でリボソーマルフレームシフトが起こり発現します。そのため、PA-XはN末端にPAとの共通領域を、C末端にPA-X特異的な領域をもちます。
PA-XはN末端にあるエンドヌクレアーゼ活性部位を介しmRNAの発現抑制を引き起こし、宿主およびウイルス蛋白質の発現を抑制します。この活性によりPA-Xは宿主の抗ウイルス応答に必須なIFN-βの発現や抗ウイルス抗体の産出を抑制し、宿主によるウイルスクリアランスを阻害します。
本研究では、N末端のエンドヌクレアーゼのみならず、C末端の6つの塩基性アミノ酸がPA-Xの発現抑制活性に重要であることを明らかにしました。本研究で得られたこれらの知見は、ウイルス蛋白質の作用機序を明らかにするための重要な手がかりとなることが期待されます。
(Nat Commun. 2015 Mar 25;6:6600)
概要
インフルエンザウイルス本来の病原性を保ち、かつ挿入した蛍光たんぱく質の発現をほぼ完全に維持できるウイルス株「Color-flu」を樹立することに成功しました。Color-fluは、蛍光たんぱく質(eCFP, eGFP, Venus, mCherry)を利用して感染細胞を光らせるので、生体内でウイルス感染が広がる様子や宿主応答をさまざまな手法で画像分析することが可能になります。本研究では、2光子レーザー顕微鏡を用いて、マウスの肺組織におけるウイルス感染細胞とマクロファージのタイムラプス撮影に初めて成功し、インフルエンザウイルスの感染により起こる免疫応答を観察することができました。さらに、Venusを発現する高病原性鳥インフルエンザウイルス(H5N1)を作製し、マウスの肺での感染の広がり方を高病原性ウイルスとインフルエンザウイルス(PR8株)とで比較することができました。Color-fluは、ウイルス病原性の解明、ウイルスに対する生体防御や気道炎症のメカニズムの解明に役立ち、さらにはワクチンや薬剤開発まで幅広く利用されることが期待されます。
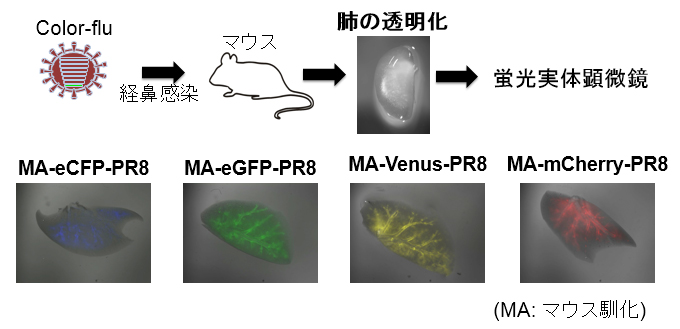
(Journal of Virology 2014 Sep;88(22):13410-7)
概要
肺炎球菌は、肺炎や中耳炎、髄膜炎などの原因となる病原体です。また肺炎球菌は、インフルエンザ罹患後の細菌性二次感染を引き起こすことが知られておりますが、インフルエンザウイルスと肺炎球菌の両方を同時に防御できるワクチンは開発されておりませんでした。
そこで今回私たちは、インフルエンザウイルスを人工合成するリバースジェネティクス法を用いて、肺炎球菌の抗原タンパク質を発現する非増殖型組み換えインフルエンザウイルスを作製いたしました(HA-KO/PspAウイルス)。
この組み換えウイルスを経鼻投与されたマウスでは、呼吸器においてインフルエンザウイルスだけでなく肺炎球菌抗原に対する抗体も誘導されることが確認され、致死的な肺炎球菌感染や、鼻腔内の菌の定着も予防できることが明らかとなりました。
今回開発された組み換えウイルスは、様々な呼吸器感染症に対するワクチンのプラットホームになることが期待されます。

概要
2013年3月下旬以降、中国において、H7N9鳥インフルエンザウイルス(H7N9ウイルス)のヒトへの感染例が報告されており、死亡例や重症化した例が多数確認されています。一般に、H7N9ウイルスは、季節性のインフルエンザウイルスとは異なり、簡単にはヒトへの感染を起こしません。そこで我々は、中国の患者から分離されたH7N9ウイルスに関する性状解析を行いました。その結果、本ウイルスが哺乳類でよく増殖すること、フェレットの間で限定的ながらも空気伝播することが分かりました(図1)。また、H7N9ウイルスに対して、ヒトが免疫を持たないことや、現在臨床で用いられているノイラミニダーゼ阻害剤の効果が低いことが明らかとなりました(図2)。従って、H7N9ウイルスによるパンデミックが起これば、甚大な被害をもたらす可能性が高いと予想されます。今回明らかにされたH7N9ウイルスの性状は、治療方法やワクチン開発、新規抗ウイルス薬の開発を含めた今後の対策を考える上で、重要な知見となります。
鳥から分離されたH7N9ウイルスと今回ヒトから分離されたH7N9ウイルスを用いて、フェレットにおけるウイルスの伝播実験を行いました。鳥から分離されたH7N9ウイルスはフェレット間での伝播は見られませんでしたが、ヒトから分離されたH7N9ウイルスはフェレット3匹中1匹においてウイルスの伝播が確認されました。
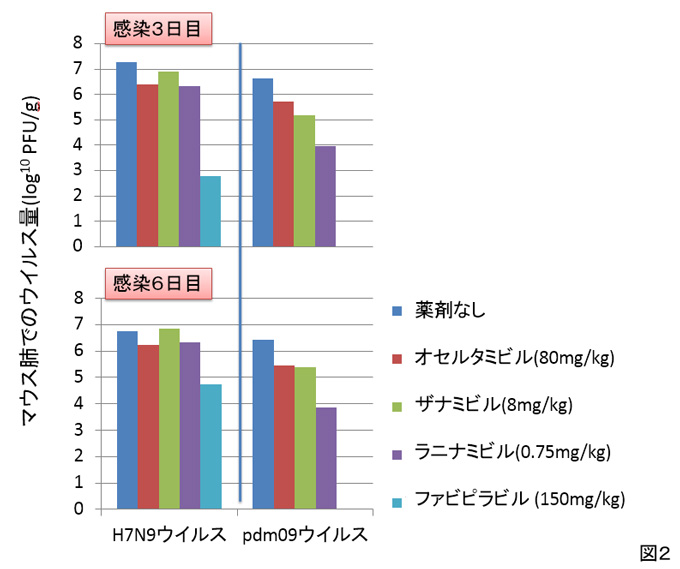
H7N9ウイルス、および、2009年のH1N1パンデミックウイルス(pdm09ウイルス)をそれぞれマウスに感染させ、2時間後に抗インフルエンザ薬を投与し、感染3日目および6日目のマウスの肺におけるウイルス量を測定しました。H7N9ウイルスは、既存のノイラミニダーゼ阻害薬であるオセルタミビル、ザナミビル、およびラニナミビルに対して低い感受性を示しましたが、未認可のRNAポリメラーゼ阻害剤であるファビピラビルには感受性を示しました。
(Journal of Virology 2013 Dec;87(23):12879-84)
概要
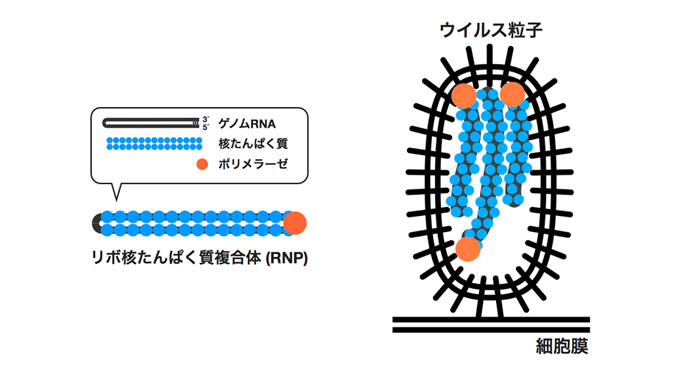
インフルエンザウイルスのゲノムRNAは8本に分節化しています。それぞれのRNA分節は、ウイルスの核たんぱく質とウイルスRNA依存的RNAポリメラーゼと共にリボ核酸たんぱく質複合体 (RNP) を形成し、ウイルス粒子内に取り込まれています (左図)。
これまで、RNPがどのような方向でウイルス粒子内に取り込まれているかは判っていませんでした。本研究では、様々な電子顕微鏡法を用いた観察によって、ウイルス粒子内には異なる方向をとったRNPが混在していることを明らかにしました (右図)。
これによって、インフルエンザウイルスの今まで知られていなかったゲノム取り込み機構の一端が明らかになったと言えます。
(Journal of Virology 2013 Jul ;87(14):7874-81)
概要
我々は、呼吸器粘膜において抗原特異的抗体を誘導できるPB2欠損非増殖型インフルエンザウイルスを作出し、その有効性を明らかにしてきました(Ozawa et al. JGV 2011, Victor et al. JV 2012)。このウイルスはPB2を欠損させた部位に外来遺伝子を搭載することで多価ワクチンへの応用が期待できます。そこで、欠損部位に、2009年にパンデミックを起こしたウイルス(pdmH1N1)およびH5N1高病原性鳥インフルエンザウイルスのHAをそれぞれ搭載したウイルスを作出し、二価ワクチンとしての効果を検証しました。
その結果、今回開発した二価ワクチンを投与したマウスでは、抗原性の異なる2つのウイルス株に対する特異的抗体が誘導されており、さらに、致死量の野生型ウイルスを感染させても100%生残しました。
以上の結果から、 PB2欠損非増殖型インフルエンザウイルスは多価ワクチンの新たなプラットフォームになることが示唆されました。
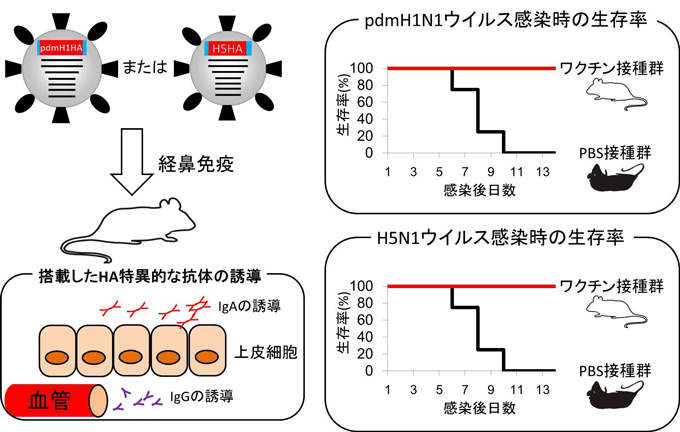
(Journal of Virology 2013 Aug;87(16):8862-9)
概要
宿主細胞に侵入後、エボラウイルスがどのような宿主蛋白質を利用してウイルスゲノムの転写・複製を行うのかは殆ど分かっていませんでした。そこで私たちは、ウイルスポリメラーゼ蛋白質(EBOL)と相互作用することでウイルスゲノムの転写・複製に関わる宿主蛋白質を探索し、DNA topoisomerase 1 (TOP1)を同定しました。本研究により、通常は核内に局在するTOP1が、ウイルスゲノムの転写・複製の場である細胞質でEBOLと相互作用すること、また、TOP1が持つ核酸切断能および核酸再結合能がウイルスゲノムの転写・複製効率に重要である事を明らかにしました。
B) EBOLとTOP1の細胞内局在。細胞質で共局在する。緑はEBOL、赤はTOP1を表す。
(Journal of Virology 2013 Mar;87(5):2455-62)
概要
A型インフルエンザウイルスのゲノムRNAは8本に分節化されています。これまで、8種類のRNA分節に合計10種類のウイルス蛋白質がコードされていると考えられてきましたが、近年、PB1-F2やPB1-N40、PA-Xなど、新たなウイルス蛋白質が見つかっています。そこで私たちはPA遺伝子分節に着目し、新たなウイルス蛋白質の探索を行いました。その結果、PA遺伝子分節に、新たなウイルス蛋白質「PA-N155」と「PA-N182」がコードされていること、さらにPA‐N155 とPA-N182がウイルス増殖やマウスにおける病原性に関与することを明らかにしました。
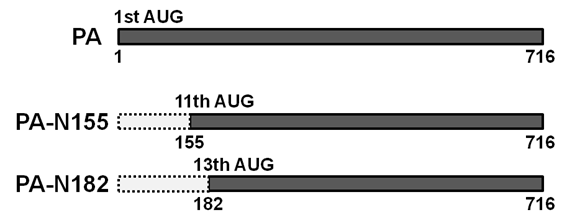
(Journal of Virology 2013 Feb;87(4):2226-33)
概要
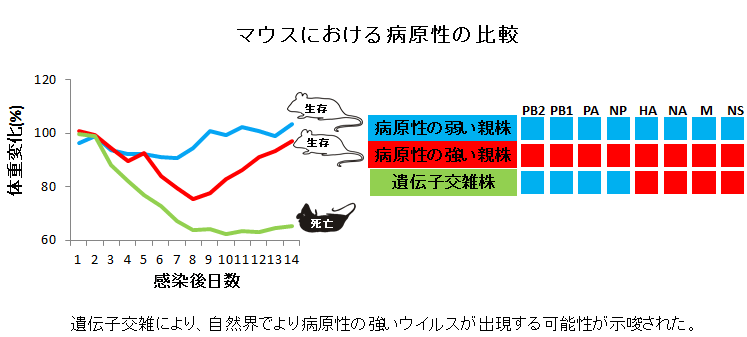
2009年インフルエンザ(以下、A(H1N1)pdm09)ウイルスによるパンデミックが発生しました。このウイルスに感染した多くの患者は軽症でしたが、一部の患者では重症化し、中には死亡する人もいました。このような予後の違いは、患者の健康状態だけでなく、ウイルス側の因子も関与していると考えられます。今回、我々は、軽症および重症患者から分離されたウイルスを用い、マウスにおける病原性を比較し、ウイルスの病原性発現因子の同定を試みました。
ウイルスの病原性発現因子を同定することは、インフルエンザウイルスの性状を知るうえで大変重要な課題であると同時に、ワクチンや抗インフルエンザ薬の開発に貢献すると考えられます。
本研究では、A(H1N1)pdm09ウイルスの複製に関わるPB2およびPB1のアミノ酸が病原性発現に関与することを明らかになり、さらに大変興味深いことに、A(H1N1)pdm09ウイルス間の遺伝子組換えにより病原性の強いウイルスが出現する可能性が示唆されました。
概要
安全で呼吸器粘膜にウイルス特異的な抗体を誘導できる新しいワクチンとして、我々は非増殖型ウイルスを基盤としたワクチン開発を行っている。
今回我々は非増殖型ウイルスとして開裂欠損型HAウイルスを作製し、マウスを用いてワクチンとしての性状解析を行った。その結果、開裂欠損型HAウイルスは効率的に液性及び細胞性免疫を誘導することが明らかとなった。また開裂欠損型HAウイルスを投与したマウスは、致死量の野生型ウイルスを感染させても100%生き残った。
以上の成績より、開裂欠損型HAウイルスは、高い免疫誘導能及び防御能を有した優れたワクチンであることが示された。
(Proc Natl Acad Sci U S A. 2012 Mar 20;109(12):4615-20)
概要
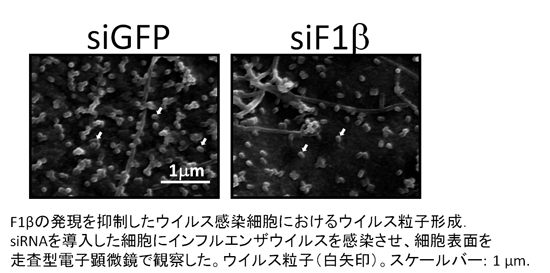
ウイルスは自らが持つ蛋白質だけでは自己複製出来ないため、増殖には我々の細胞(宿主)の分子が必要です。ウイルス増殖に関わる細胞の分子を明らかにすることは『ウイルスが我々の細胞の中でどうやって増殖しているのか?』という疑問に答えるために必要な、パズルのピース集めのようなものです。本研究で我々は、人間の細胞内でインフルエンザウイルスの蛋白質(NS2)がくっつき、ウイルスが細胞の外に放出される際に必要な分子(F1β)を見つけました。今後、宿主の分子を標的とした抗ウイルス薬の開発が可能なのではないかと期待しています。