
腸管における病原細菌(赤痢菌)と宿主自然免疫のあらたな攻防を解明:新規病原因子の立体構造とその標的宿主因子の発見
腸管における病原細菌(赤痢菌)と宿主自然免疫のあらたな攻防を解明:新規病原因子の立体構造とその標的宿主因子の発見
毎年約1500万人が感染症で命を失われ、そのなかの約200万人は腸管感染症が原因である。腸管は無数の微生物に常に暴露されているが、微生物の侵入から生体を守るために自然免疫を中心とする堅固な防御システムが幾重にも備えられている。一方、赤痢菌やその仲間である病原性大腸菌(O157等)等の腸管病原細菌は、それら防御システムを巧みに回避して生体へ侵入する高度に進化した機能を備えている。病原細菌の腸粘膜への侵入に対する自然免疫応答と、それを病原細菌がどのようにして回避するか、そのメカニズムは全く不明であった。
今回、東京大学医科学研究所の笹川千尋教授と真田真人研究員らは、兵庫県立大学の水島恒裕教授および大型放射光施設(SPring-8)との共同研究により、赤痢菌をモデルにして、(i) 粘膜上皮に対する病原体の侵入認識機構。(ii) この宿主認識・防御に対抗する赤痢菌の手段 を明らかにした。具体的には、(i) 細菌が細胞侵入するときに形成される葉状突起(ラッフル膜)を危険信号として認識する仕組みとして、「葉状突起に局在するジアシルグリセロール(DAG)-TRAF6-NF-κBに依存した炎症シグナル経路が重要である」ことを明らかにした。(ii) 赤痢菌の対抗手段として菌から分泌されるOspIを同定し、そのタンパク質の立体構造(図2参照)および生化学的性状を解明した。(i)と(ii)の結果から、OspIは、上述の炎症シグナル経路の制御に重要なTRAF6の活性化に必要なUBC13に結合性して、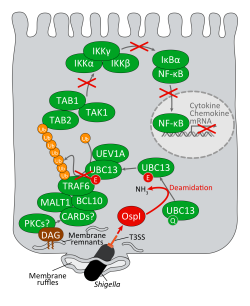
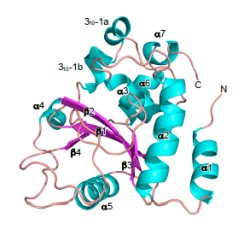 UBC13の100番目のグルタミンを脱アミド化する「新規な脱アミド化酵素である」ことを発見した(図1参照)。本研究により、感染初期の病原体に対する粘膜上皮の新規な防御機構と、それに対抗する病原体側のあらたな戦術が解明され、これを標的に創薬やワクチン開発への応用が期待される。
UBC13の100番目のグルタミンを脱アミド化する「新規な脱アミド化酵素である」ことを発見した(図1参照)。本研究により、感染初期の病原体に対する粘膜上皮の新規な防御機構と、それに対抗する病原体側のあらたな戦術が解明され、これを標的に創薬やワクチン開発への応用が期待される。