
| 発表のポイント |
|---|
|
概要
東京大学医科学研究所RNA制御学分野の稲田利文教授、松尾芳隆准教授、友松翔太大学院生(研究当時)らの研究グループは、同研究所タンパク質代謝制御分野の佐伯泰教授、富田拓哉助教、星薬科大学の大竹史明准教授らと共同で、リボソームの異常渋滞の解消に必須なリボソームタンパク質uS10のユビキチン修飾について、高次構造とその機能、および制御メカニズムを世界で初めて解明しました。先行研究より、uS10にK63鎖が形成されることでリボソームの異常渋滞が解消されることが知られていました。本研究では生化学的な手法を用いることで、K63鎖の他に、K48鎖やK48/K63混合鎖が形成されていることを新たに発見しました。先行研究と合致してK63鎖は異常渋滞を解消する一方で、K48鎖とK48/K63混合鎖は解消しないことが明らかとなりました。また、遺伝学的スクリーニングにより同定したuS10に対する脱ユビキチン化酵素のUbp2とUbp3が、ユビキチン修飾を編集し、リボソームの異常渋滞の解消に持続的な活性を与えることを明らかにしました。本成果は、リボソームの異常渋滞が原因とされる神経変性疾患などの発症機序の理解や新規治療戦略の開発に繋がることが期待されます。
本研究成果は2025年9月17日付で、国際科学雑誌「THE EMBO JOURNAL」オンライン版で公開されました。
発表内容
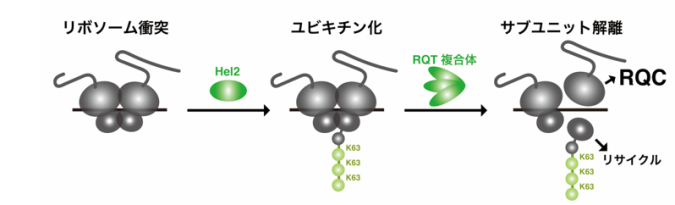
DNAにコードされた遺伝情報の多くは、転写、翻訳を経て合成されたタンパク質が機能することで発現します。遺伝子発現は生命現象の根幹であるため、さまざまな品質管理によって、その正確性が保証されています。タンパク質合成反応である翻訳では、異常な翻訳停滞を識別し、途中まで合成された新生ペプチド鎖を分解する品質管理RQC(Ribosome-associated Quality Control:注1)が働いています。この品質管理では、リボソーム(注2)の衝突を認識したユビキチン付加酵素のHel2が、リボソームタンパク質uS10に対してユビキチン修飾(注3)を付加し、異常を警告します。デコーダー分子のRQT複合体(注4)はこれを感知し、停滞したリボソームをサブユニット解離することで、異常な翻訳停滞が解消されます(図1)。この分子メカニズムは真核生物において広く保存されており、その重要性が伺えます。
ユビキチン修飾は、多様な生命現象に関与する重要な翻訳後修飾です。ポリユビキチン修飾には8種類の連結型が存在し、代表的なものとしてK48型はプロテアソーム分解、K63型は非プロテアソーム分解に関わることが知られています。近年、複数の連結型が組み合わさった混合鎖が発見され、ユビキチン修飾の高次構造が、その機能を決定づけることが明らかとなってきました。そのため、ユビキチン修飾にコードされた機能暗号の解読は、生命現象を理解する上で重要な研究として注目を集めています。
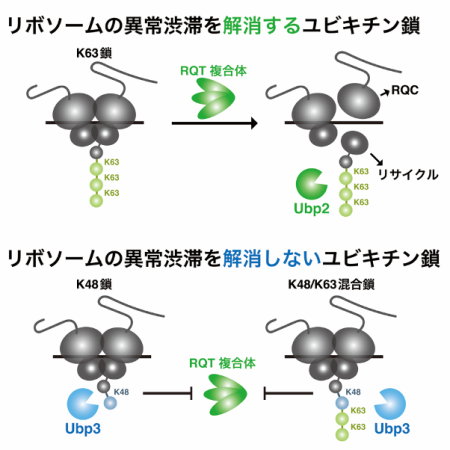
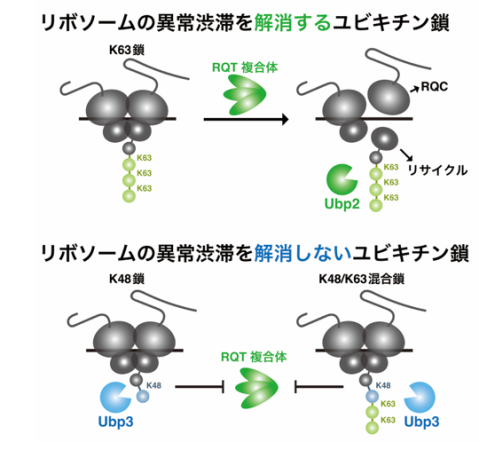
先行研究より、uS10のK63型ユビキチン修飾が異常な翻訳停滞の解消に必須であることが明らかとなっていました(図1)。K48型ユビキチン修飾もuS10に形成されている可能性が示唆されていましたが、その機能や制御メカニズムについては不明な点が多く残されていました。
本研究では、生化学的解析により、uS10のユビキチン修飾の高次構造には、K63鎖、K48鎖、根本にK48型、遠位にK63型が形成されたK48/K63混合鎖の3つのタイプが存在することを明らかにしました。RQT複合体のユビキチン鎖への選択制から、K63鎖は異常翻訳停滞の解消に働くのに対し、K48鎖とK48/K63混合鎖は異常翻訳停滞の解消に働かないことが明らかとなりました(図2)。
また、遺伝学的スクリーニングにより、uS10に対する脱ユビキチン化酵素(注5)として同定したUbp2とUbp3について機能解析を実施しました。Ubp2は解離した40Sサブユニット上でK63鎖を除去する一方で、Ubp3は翻訳途中のリボソームに形成されたK48鎖とK48/K63混合鎖を除去していることが明らかとなり、脱ユビキチン化酵素の基質特異性に明確な違いが判明しました。また、これらの脱ユビキチン化酵素が、uS10のユビキチン修飾を編集することで、異常翻訳停滞の解消を持続的に活性化することを明らかにしました(図2)。
近年、異常な翻訳停滞の蓄積が、アポトーシス(注6)や自己免疫応答(注7)などのさまざまな細胞応答を誘導することが明らかとなっています。本研究は、異常な翻訳停滞の解消の狼煙として機能するuS10のユビキチン修飾について、高次構造とその機能、及び、制御メカニズムを明らかにしました。本成果は、品質管理機構の破綻が原因とされる神経変性疾患などの発症機序の理解や新規治療戦略の開発に繋がることが期待されます。
発表者・研究者等情報
東京大学医科学研究所 基礎医科学部門 RNA制御学分野
稲田 利文 教授
兼:同大学大学院理学研究科 生物科学専攻
松尾 芳隆 准教授
星薬科大学 先端生命科学研究所
友松 翔太 特任助教
研究当時:東京大学大学院薬学系研究科薬科学専攻 博士課程
論文情報
雑誌名:THE EMBO JOURNAL題 名:Polyubiquitin architecture editing on collided ribosomes maintains persistent RQC activity
著者名: Shota Tomomatsu, Yoshitaka Matsuo*, Fumiaki Ohtake, Takuya Tomita, Yasushi Saeki, Toshifumi Inada* (*責任著者)
DOI: 10.1038/s44318-025-00568-0
URL: https://www.embopress.org/doi/epdf/10.1038/s44318-025-00568-0
研究助成
本研究は、日本医療研究開発機構(AMED-CREST 課題番号:20gm1110010h0002:稲田利文)、「ワクチン開発のための世界トップレベル研究開発拠点の形成事業(UTOPIA, JP223fa627001:稲田利文)、武田科学振興財団研究助成(稲田利文)、科学技術振興機構(JST)戦略的創造研究推進事業さきがけ「細胞の動的高次構造体」領域(課題番号:JPMJPR21EE:松尾芳隆)、「加齢による生態変容の基盤的な理解」領域(課題番号:JPMJPR2488:松尾芳隆)、日本学術振興会科学研究費助成事業(課題番号: 19H05281、21H05277、22H00401、25H00007、稲田利文; 25H01442, 25K02221 松尾芳隆; 18H05498佐伯泰 & 大竹史明 )、などの支援を受けて行われました。
用語解説
(注1)品質管理機構RQCRQC(Ribosome-associated Quality Control)は、異常な翻訳停滞に対し、停滞したリボソームをmRNA上から解離すると共に不良タンパク質を分解へ導く機構である。
(注2)リボソーム
メッセンジャーRNA(mRNA)の持つ遺伝情報に従ってタンパク質を合成する装置。複数のタンパク質とRNAから構成される巨大な複合体である。
(注3)ユビキチン修飾
ユビキチンは76アミノ酸からなる低分子タンパク質である。ユビキチンが他のタンパク質のリジン残基に共有結合で付加されると、タンパク質分解やシグナル伝達などを制御する。
(注4)RQT複合体
RNAヘリカーゼと2種類のユビキチン結合タンパク質からなる複合体。翻訳停滞したリボソームを解離する活性をもつ。
(注5)脱ユビキチン化酵素
ユビキチン修飾を切断する酵素。特定の連結型のみ切断するものや、特性を示さないものもある。出芽酵母では約20種類、ヒトでは100種類あるとされている。
(注6)アポトーシス
多細胞生物の細胞で増殖制御機構として管理・調節された、能動的な細胞死であり、プログラム細胞死とも呼ばれる。
(注7)自然免疫応答
病原体感染初期の感染防御を担う免疫機構。
問合せ先
〈研究に関する問合せ〉東京大学医科学研究所 基礎医科学部門 RNA制御学分野
教授 稲田 利文(いなだ としふみ)
〈報道に関する問合せ〉
東京大学医科学研究所 プロジェクトコーディネーター室(広報)
https://www.ims.u-tokyo.ac.jp/