
| 発表のポイント |
|---|
|
概要
東京大学医科学研究所RNA制御学分野/同大学大学院理学系研究科生物科学専攻の稲田利文教授と同大学大学院理学系研究科生物科学専攻の佐藤二千翔大学院生らによる研究グループは、小胞体ストレス(注1)条件下における、リボソーム(注2)のユビキチン化(注3)に起因する翻訳制御の仕組みを明らかにしました。本研究では出芽酵母のE3ユビキチン化酵素欠損株ライブラリーを用いたスクリーニングにより、小胞体ストレス応答(UPR)に関わる因子としてGrr1を新たに同定しました。またその後の詳細な解析により、「分解因子Grr1が脱ユビキチン化酵素を分解することで間接的にリボソームユビキチン化レベルを上昇させ、転写因子(注4)HAC1 mRNAの翻訳を促進する」という分子機構を明らかにしました。先行研究と比較して、本研究は小胞体ストレス応答についてリボソームユビキチン化に起因する翻訳制御という新たな切り口から解析を行った点で新規性があり、この研究成果は今後新たな治療法や創薬への応用に役立つことが期待されます。本研究成果は2025年3月4日、「Nature Communications」にオンライン掲載されました。
発表内容
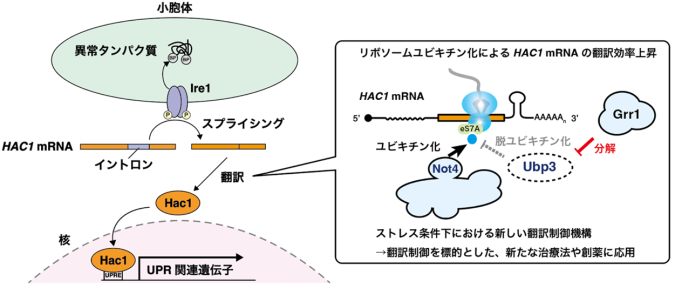
出芽酵母におけるUPRでは、転写因子Hac1が主要な役割を担います。通常条件下ではHAC1 mRNAはイントロン(注5)と5’UTR(注6)が塩基対形成することで翻訳が強く抑制されていますが、小胞体ストレス条件下ではセンサータンパク質Ire1によるスプライシングによってイントロンが除去されることで活発に翻訳されるようになります(図1)。
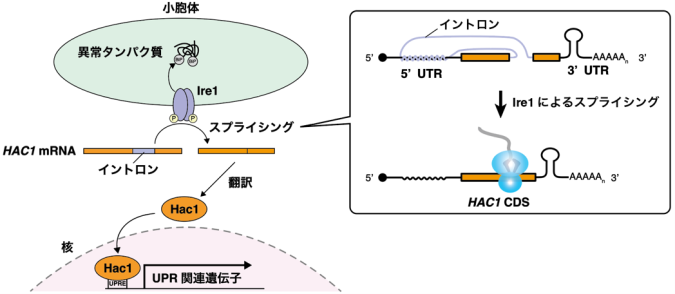
転写因子Hac1の発現は厳密に制御されており、通常条件下ではイントロンと5’UTRの相互作用によって翻訳が強く抑制されている一方、小胞体ストレス条件下ではスプライシングによってイントロンが除去されることで活発に翻訳されるようになる。
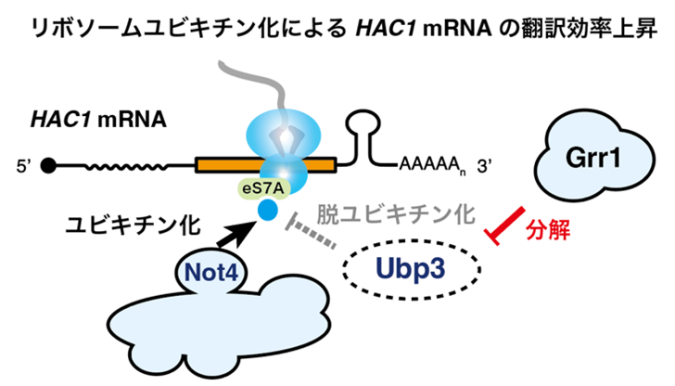
これまでの先行研究では主にスプライシングの分子機構について焦点が当てられていましたが、本研究チームはスプライシング後のHAC1 mRNAの翻訳に焦点を当て研究を行いました。本研究チームの以前の先行研究により、「E3ユビキチン化酵素Not4によるリボソームタンパク質eS7Aのユビキチン化が、効率的なHAC1 mRNAの翻訳に必須であること」、「小胞体ストレス条件下ではeS7Aの脱ユビキチン化酵素Ubp3の発現が減少すること」が判明しており、本研究では「Ubp3の発現減少を担う因子」と「HAC1 mRNAが持つ、eS7Aユビキチン化による翻訳制御の標的となる領域」について明らかにすることを目的としました(図2)。
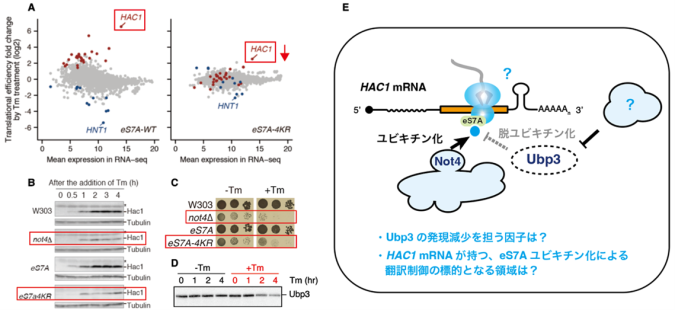
(A)ユビキチン化が起こらないeS7A-4KR株では、HAC1の翻訳効率が減少する。
(B,C) not4欠損株及びeS7A-4KR株では、Hac1発現が低下し、ツニカマイシン(Tm)が誘導する小胞体ストレスに感受性を示す。
(D)脱ユビキチン化酵素のUbp3は、小胞体ストレス条件下で発現が減少する。
(E)先行研究により考案されたモデル図と、本研究で取り組んだ疑問点。
まず、E3ユビキチン化酵素欠損株ライブラリーを用いたスクリーニングにより、UPRに関わる因子としてGrr1を新たに同定しました。その後のUbp3の安定性やeS7Aユビキチン化レベルの評価、リボソーム・プロファイリング(注7)による翻訳動態の詳細な解析により、「Grr1が脱ユビキチン化酵素Ubp3を分解することで間接的にリボソームユビキチン化レベルを上昇させ、転写因子HAC1 mRNAの翻訳を促進する」という分子機構を明らかにしました(図3)。
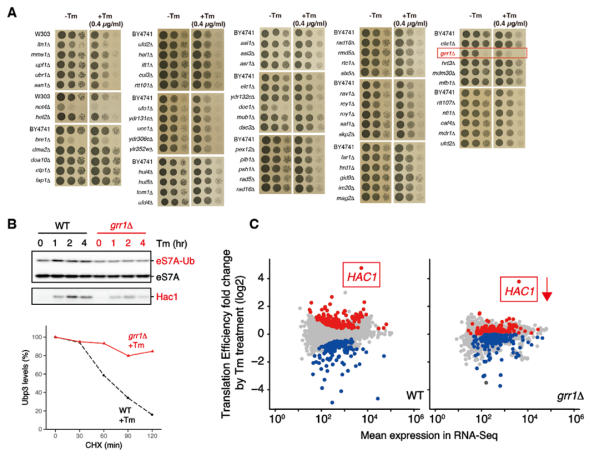
(A,B) grr1欠損株はツニカマイシン(Tm)が誘導する小胞体ストレスに感受性を示し、eS7Aユビキチン化レベルの減少、Hac1発現の低下、Ubp3の安定化がみられる。
(C) grr1欠損株では、HAC1の翻訳効率が減少する。
また、種々のHAC1レポーターを用いた解析により、HAC1 mRNAは自身のコーディング領域CDS(注8)内に、eS7Aユビキチン化による翻訳制御の標的となる配列を持つことを発見しました。一方で、HAC1 mRNAのUTRはHac1の発現量上昇には寄与するものの、eS7Aユビキチン化による翻訳制御には関与しないことが明らかとなりました(図4)。
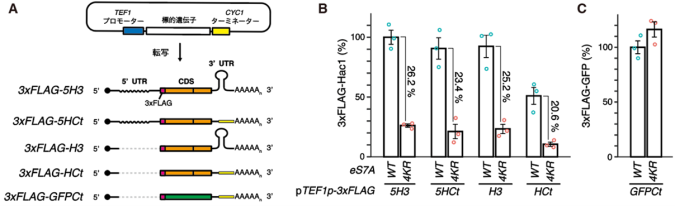
(A)用いたレポーターの模式図。
(B,C)HAC1 mRNAはCDSのみ(HCt)でも、eS7Aユビキチン化による翻訳制御を受ける。
小胞体ストレスは糖尿病、アルツハイマー病、 がんなどの疾患を誘導することが知られており、また小胞体ストレス治療薬として、化学シャペロン(注9)の 4-PBAなどが存在します。一方で、翻訳に焦点を当てた小胞体ストレス治療薬は開発されていません。またUPRにはeIF2αリン酸化(注10)による翻訳抑制機構が存在しますが、本研究はそれとは異なる新しい経路のリボソームユビキチン化に起因する翻訳制御機構を報告しました。 得られた知見は、ストレス状態や疾患全体での翻訳動態を理解する足掛かりとなり、新たな治療法や創薬に応用できると期待されます(図5)。
効率的なHAC1 mRNAの翻訳に寄与する
発表者・研究者等情報
東京大学医科学研究所 基礎医科学部門 RNA制御学分野/東京大学大学院理学系研究科 生物科学専攻稲田 利文 教授
東京大学大学院理学系研究科 生物科学専攻
佐藤 二千翔 博士課程1年
論文情報
雑誌名:Nature Communications題 名:Crucial roles of Grr1 in splicing and translation of HAC1 mRNA upon unfolded stress response
著者名:Nichika Sato, Yu Nakano, Yasuko Matsuki, Shota Tomomatsu, Sihan Li, Yoshitaka Matsuo, Toshifumi Inada*(*責任著者)
DOI: 10.1038/s41467-025-57360-1
URL: https://www.nature.com/articles/s41467-025-57360-1
研究助成
本研究は、日本医療研究開発機構(AMED-CREST 課題番号:20gm1110010、研究代表者:稲田利文)、日本学術振興会科学研究費助成事業(課題番号:JP18H03977, JP19H05281、稲田利文;21H00267, 21H05710, 22H02606、松尾芳隆)、科学技術振興機構(JST)さきがけ(課題番号:JPMJPR21EE、研究代表者:松尾芳隆)などの支援を受けて行われました。
用語解説
(注1)小胞体ストレスタンパク質の適切な折りたたみを行う場である小胞体に、さまざまなストレスによってその機能が阻害されることで、折りたたみ不全の異常タンパク質が蓄積した状態をいう。小胞体ストレス状態を改善するために、生体は小胞体ストレス応答(UPR)を発動する。UPRは真核生物全てに保存されており、どの生物種でも異常タンパク質を減らすことで小胞体の負荷を軽減しようとする点で共通している。
(注2)リボソーム
大小二つのサブユニットからなり、翻訳(mRNAにコードされた遺伝暗号であるコドン配列に基づいてアミノ酸を連結させていき、タンパク質を合成する過程)を行う。リボソームタンパク質とリボソームRNAから構成されており、真核生物・原核生物のどちらにも存在するが、その大きさは異なっている。
(注3)ユビキチン化
基質となるタンパク質に、76アミノ酸からなる小さなタンパク質であるユビキチンが付加される現象。ユビキチン化は三段階の反応からなり、まずATPによりE1ユビキチン活性化酵素がユビキチンを活性化する。活性化されたユビキチンはE2ユビキチン結合酵素へと転移し、基質へはE3ユビキチン化酵素が結合させる。ユビキチンが結合する場所や様式の違いによって、その後の基質タンパク質の運命やシグナル伝達が変化する。また基質に付加されたユビキチンは、脱ユビキチン化酵素によって取り除かれ、再利用される。
(注4)転写因子
特定のDNA配列に結合し、転写(DNA配列に基づいてRNAを合成する過程)を調節する因子。本研究対象であるHac1は、UPREと呼ばれる配列に結合することで、下流のUPR関連遺伝子の転写を促進する。
(注5)イントロン
転写直後のRNAには含まれているものの、最終的な転写産物からは除去される配列。イントロンを除去する過程をスプライシングという。
(注6)UTR
非翻訳領域(Untranslated region)といい、タンパク質には翻訳されない領域。タンパク質に翻訳される領域の上流にあるUTRを5’UTR、下流にあるUTRを3’UTRという。
(注7)リボソーム・プロファイリング
リボソームによって覆われているmRNA断片を回収し、次世代シーケンサーで網羅的に解析する手法。mRNA上のリボソームの位置を特定したり、それぞれのmRNAでの翻訳量や翻訳速度を測定したりすることで、細胞内の翻訳動態解析が可能である。
(注8)コーディング領域(CDS: Coding sequence)
タンパク質に翻訳される領域。一方ORF(Open reading frame)は開始コドンから終止コドンまでの領域をいい、イントロンを含む。
(注9)シャペロン
タンパク質の適切な折りたたみを補助する機能を持つ。この中でも、低分子化合物であるものを化学シャペロンという。
(注10)eIF2αリン酸化
eIF2αは翻訳開始因子の一つであるeIF2のサブユニットである。さまざまなストレスによってeIF2αがリン酸化されると、新たなタンパク質の翻訳開始が抑制される。一方で、uORF(本来のCDSの5’上流に開始コドンが存在し、翻訳が始まる領域)を持つ一部の遺伝子の翻訳は活性化される。
問合せ先
〈研究に関するお問い合わせ〉東京大学医科学研究所 基礎医科学部門 RNA制御学分野
教授 稲田 利文(いなだ としふみ)
https://www.ims.u-tokyo.ac.jp/imsut/jp/lab/basicmedicalsciences/page_00154.html
〈報道に関するお問い合わせ〉
東京大学医科学研究所プロジェクトコーディネーター室(広報)
https://www.ims.u-tokyo.ac.jp/
東京大学大学院理学系研究科・理学部 広報室
https://www.s.u-tokyo.ac.jp/ja/