研究内容
高等生物の発生と生殖は、多様な細胞が時間的・空間的に協調することで成立する精密な生命現象です。私たちの研究室では、ゲノム編集、遺伝子改変マウス・ラット作製、多能性幹細胞、単一細胞解析などを組み合わせ、発生・生殖を支える分子機構を個体レベルで理解することを目指しています。特に、精子形成、生殖細胞を支える精巣組織環境、ゲノム編集によるモデル動物作製技術、そしてヒト疾患や種差を解析するための遺伝子全長ヒト化マウス作製技術の開発に取り組んでいます。
精子形成の制御の解明
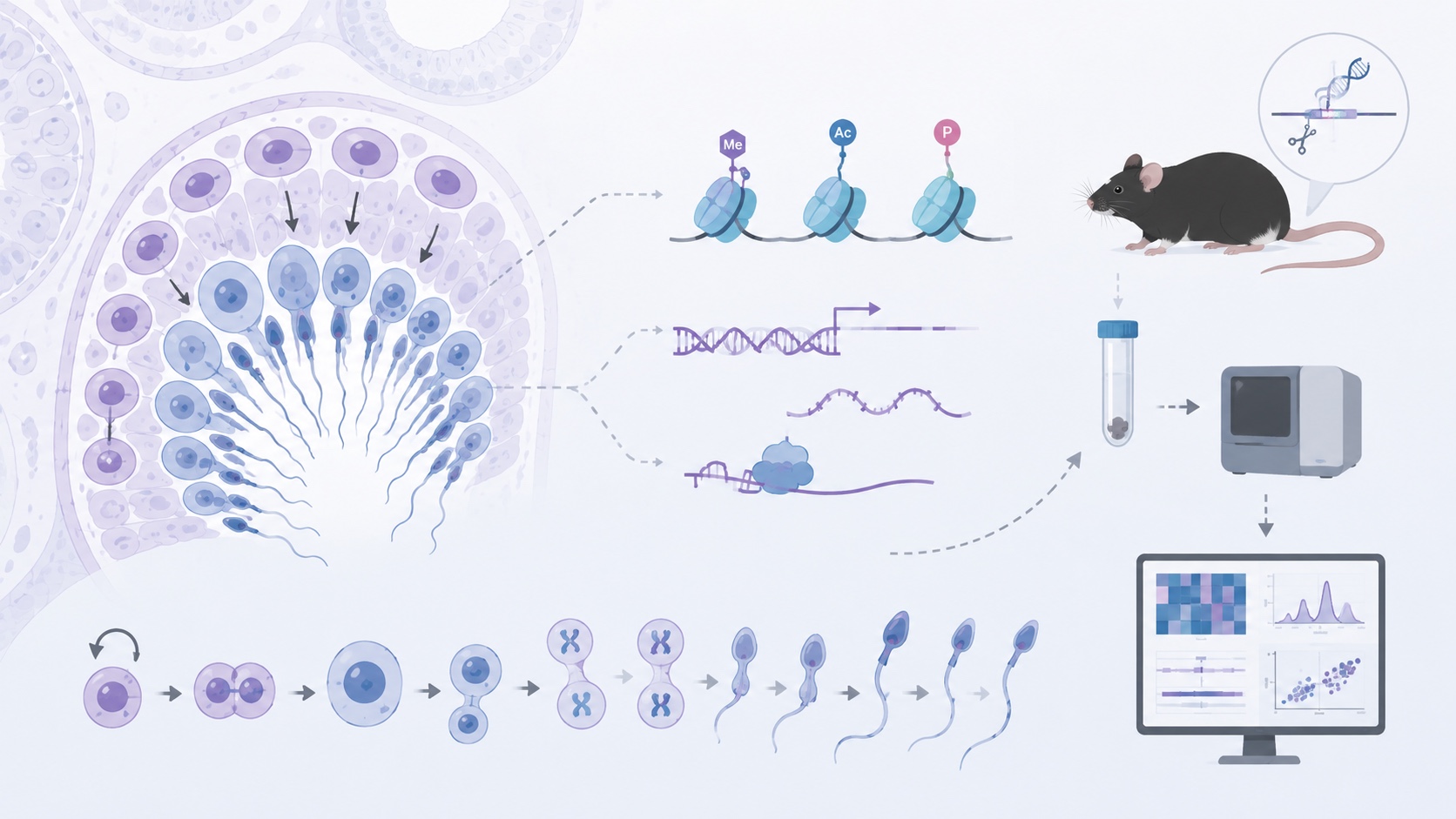
生殖細胞は、遺伝情報を次世代へ受け渡すために高度に特殊化した細胞です。精子形成では、精原細胞の自己複製と分化、減数分裂、精子細胞の形態形成が、長い時間軸の中で厳密に制御されています。私たちは、遺伝子改変マウスや分子生物学的解析を用いて、精子形成を制御する遺伝子機能、エピゲノム制御、RNA制御の仕組みを明らかにしようとしています。これまでに、ヒストン修飾制御因子が精原細胞の増殖や加齢に伴う精子形成能力の維持に関わることを見出しており、生殖細胞が長期にわたり機能を保つ分子基盤の解明を進めています。
精子形成を支える精巣体細胞の機能的役割の解明
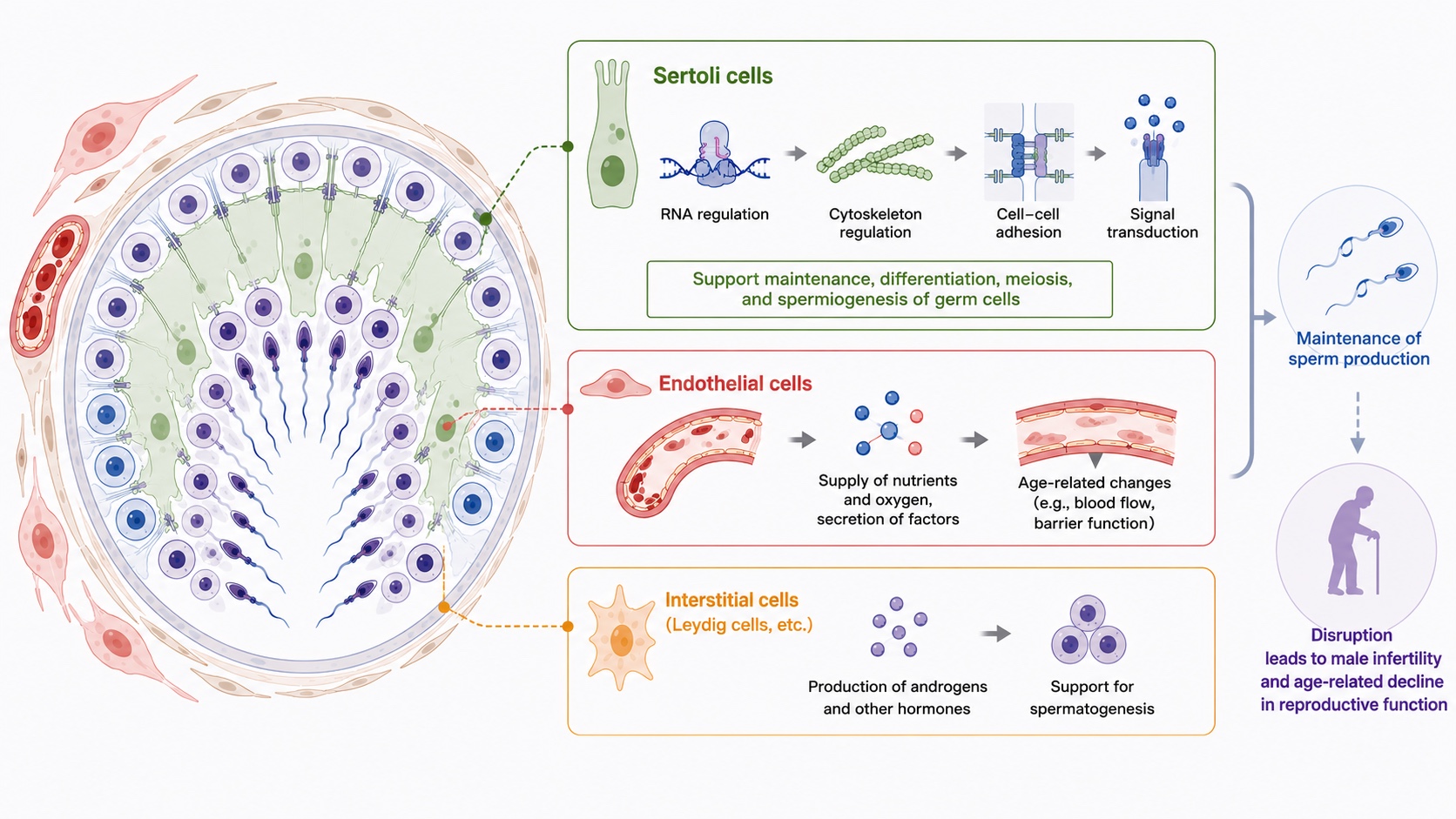
精子形成は、生殖細胞だけで完結する現象ではありません。精巣内では、セルトリ細胞、内皮細胞、間質細胞などの体細胞が、生殖細胞の維持、分化、減数分裂、形態形成を支えています。私たちは、精巣体細胞がどのように精子形成を制御し、その破綻が男性不妊や加齢に伴う生殖機能低下につながるのかを解析しています。特に、セルトリ細胞におけるRNA制御、細胞骨格制御、細胞間接着、シグナル伝達が、精子形成を支える仕組みに着目しています。また、精巣内皮細胞の加齢変化が精子形成能力の低下に関与することを見出しており、生殖機能を支える組織環境の理解を進めています。
ゲノム編集による遺伝子改変動物作製技術の高度化
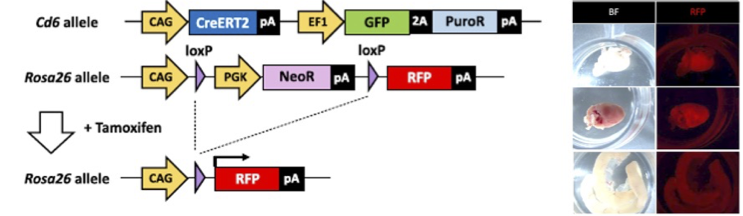
遺伝子改変動物は、遺伝子機能を個体レベルで検証するための重要な研究基盤です。私たちは、マウスをモデルとして、CRISPR/Cas9をはじめとするゲノム編集技術とES細胞を組み合わせ、ノックアウト、ノックイン、条件付き変異導入、大規模ゲノム改変を効率よく実現する技術の開発を進めています。これまでに、ES細胞における遺伝子改変効率を大きく向上させ、複数遺伝子を同時に高効率で改変できる技術を報告してきました。これらの技術を基盤として、発生・生殖研究、疾患モデル作製、遺伝子機能解析に利用できる高精度なモデル動物作製を進めています。また、マウスで確立した先端技術のラットへの応用も目指しています。
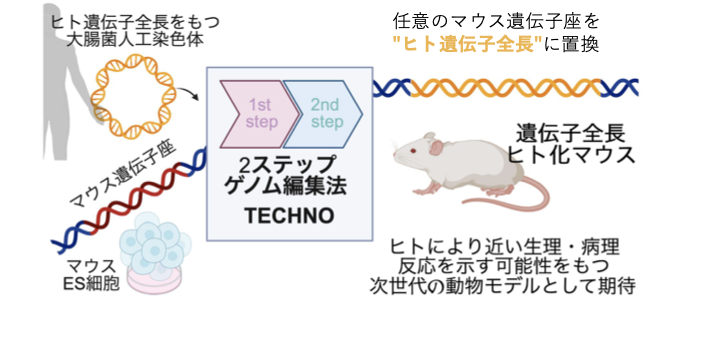
遺伝子全長ヒト化マウスによるヒト疾患・種差の理解
ヒト疾患の理解には、タンパク質コード領域だけでなく、イントロン、非翻訳領域、調節配列を含む遺伝子全体の機能を解析することが重要です。しかし、従来のマウスモデルでは、ヒト特異的な遺伝子制御、スプライシング、非コード領域の機能を個体レベルで再現することは容易ではありません。私たちは、ヒト遺伝子を周辺制御領域を含めてマウスゲノム上に再構築する遺伝子全長ヒト化マウス作製技術を開発し、ヒト疾患関連変異や種差を個体レベルで解析できる新しいモデル動物研究を進めています。