
| 慶應義塾大学大学院薬学研究科の服部きさら(博士課程修了)、同大学院丸田ひかり(博士課程在籍)、髙橋大輔准教授、長谷耕二教授(東京大学医科学研究所 共生生物学分野 教授、福島大学 食農学類附属発酵醸造研究所 特任教授 兼任)らの研究グループは、慶應義塾大学、東京大学医科学研究所、福島大学をはじめとする国内複数機関との共同研究により、食事に含まれる大豆が腸内の特定の共生細菌(Limosilactobacillus reuteri と Muribaculum intestinale)の定着を促進し、この2種の細菌が協力して小腸パイエル板※1の免疫細胞(濾胞性ヘルパーT細胞※2)を活性化することで、幅広い細菌に結合できる「多応答性IgA抗体※3」の産生を誘導することを発見しました。さらに、L. reuteri がT細胞受容体に認識される抗原として、M. intestinale が樹状細胞からのIL-1β産生を促すアジュバント(補助剤)として機能するという、2菌種の「役割分担」による協調メカニズムを解明しました。大豆食により誘導された多応答性IgAは、腸内細菌叢の恒常性維持に寄与するとともに、サルモネラ菌をはじめとする病原菌に対する感染防御能を高めることも明らかにしました。 本研究は、食事と腸内細菌が協調して粘膜免疫を形作る「食事—腸内細菌—免疫軸」の実像を分子レベルで明らかにしたものであり、将来的には、腸内細菌叢の正常化と感染症予防を両立する新たな機能性食品・プロバイオティクス戦略の開発や、離乳期の栄養に基づく予防医学への応用につながることが期待されます。 本研究成果は2026年5月20日に、国際学術誌 Immunity に掲載されました。 |
1. 本研究のポイント
2. 研究の背景
私たちの腸内には約40兆個の共生細菌が生息しており、宿主の健康維持に重要な役割を果たしています。なかでも分泌型IgA抗体※5は、腸内細菌叢の構成を制御し、宿主と共生細菌の共存関係を維持する中心的な役割を担っています。小腸のパイエル板では、濾胞ヘルパーT(Tfh)細胞が胚中心反応を駆動することで、高親和性のIgA抗体が産生されることが知られています。パイエル板のTfh細胞は無菌マウスではほとんど観察されず、腸内細菌依存的に分化することが知られていますが、どの腸内細菌が、どのようなメカニズムで、どのような食事環境下でTfh細胞の分化を誘導するのかは十分に解明されていませんでした。特に、離乳期は腸内細菌叢の構築と粘膜免疫系の成熟が同時に進行する重要な時期であり、この時期の食事が腸管免疫の長期的な応答様式にどのような影響を及ぼすかは、未解明の重要課題として残されていました。
3. 研究の内容・成果
(1)大豆がパイエル板のTfh細胞とIgA応答を増強する
離乳期のマウスに精製飼料(AIN-93G)を与えると、パイエル板のTfh細胞、胚中心B細胞、IgA陽性B細胞が著しく減少しました。一方、非精製飼料(CE-2)では正常な応答が維持されました。CE-2飼料の原材料を個別にAIN-93Gに添加してスクリーニングした結果、大豆粉の添加のみがこれらの免疫応答を回復させることを見出しました(図1)。この効果は無菌マウスでは消失し、M細胞※6欠損マウスおよびTfh細胞欠損マウスでも観察されなかったことから、大豆の免疫増強効果は腸内細菌依存的かつM細胞-Tfh細胞経路を介していることが示されました。
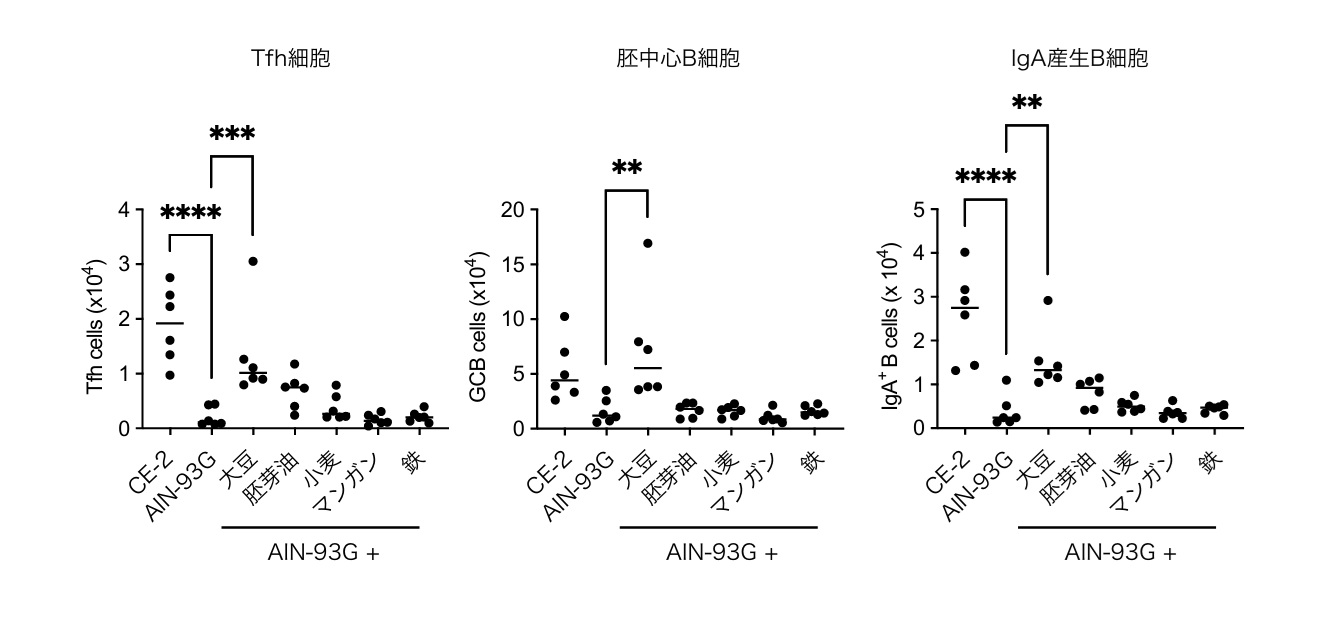
離乳期マウスに、通常飼料(CE-2)、精製飼料(AIN-93G)、またはAIN-93GにCE-2の主要原材料(大豆粉、コーン胚芽油、小麦粉、マンガン、鉄)を個別に添加した飼料を給与し、3週間後に小腸パイエル板のTfh細胞数(左)、胚中心B細胞数(GCB, 中央)、IgA産生B細胞数(右)を解析した。CE-2群に比べAIN-93G群ではいずれも顕著に減少したのに対し、AIN-93Gに大豆粉のみを添加した群で選択的に応答が回復した。
(2)大豆は L. reuteri と M. intestinale の定着を促進する
大豆食マウスの腸内細菌叢を16S rRNA解析で解析したところ、細菌叢の構成がCE-2飼料摂食マウスに近づき、特に L. reuteriと M. intestinaleの2菌種が選択的に増加し、パイエル板のTfh細胞数と正の相関を示しました。無菌マウスを用いたノトバイオート実験により、この2菌種の同時定着下でのみTfh細胞とIgA応答が誘導されることを証明しました(図2)。M. intestinaleを、飼育施設で存在量が最も豊富なF. rodentiumに置き換えた置換実験では応答が再構成されなかったことから、本協調作用は M. intestinaleに特異的であることが確認されました。
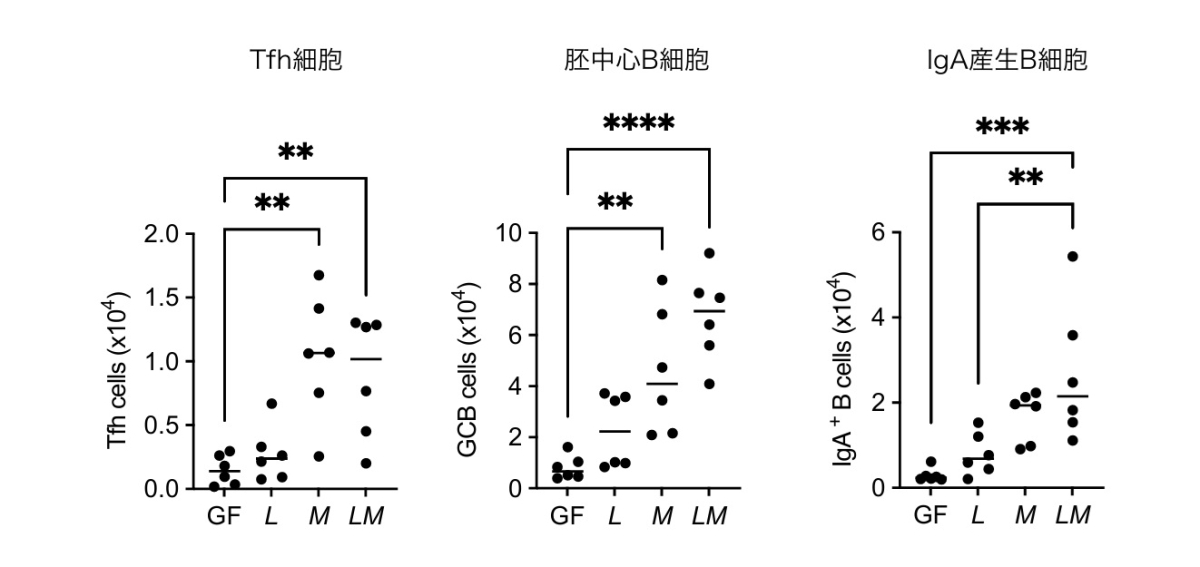
無菌(germ-free; GF)マウスに L. reuteri単独(L)、M. intestinale単独(M)、または両者を同時(LM)に経口接種してノトバイオートマウスを作製し、4週間後にパイエル板のTfh細胞、胚中心B細胞、IgA産生B細胞を解析した。GF群および単独定着群ではいずれの細胞も誘導されなかったのに対して、LM共定着群でのみすべての細胞が有意に増加し、両菌種の協調作用が必要であることが示された。
(3)L. reuteriが抗原、M. intestinaleがアジュバントとして協働する
Tfhハイブリドーマ81クローンの抗原特異性を解析した結果、全クローンの10%以上がL. reuteriに反応性を示したことから、L. reuteriがパイエル板Tfhレパートリーの主要な抗原源として機能していることを明らかにしました。一方、M. intestinaleは骨髄由来樹状細胞からのIL-1β産生を強力に促進し、アジュバントとして機能していました(図3)。パイエル板のシングルセルRNA-seq解析でも、大豆食群のcDC(Conventional Dendritic Cell)のサブセットでIL-1β遺伝子の発現上昇が確認され、2菌種の「抗原-アジュバント」役割分担が分子レベルで裏付けられました。
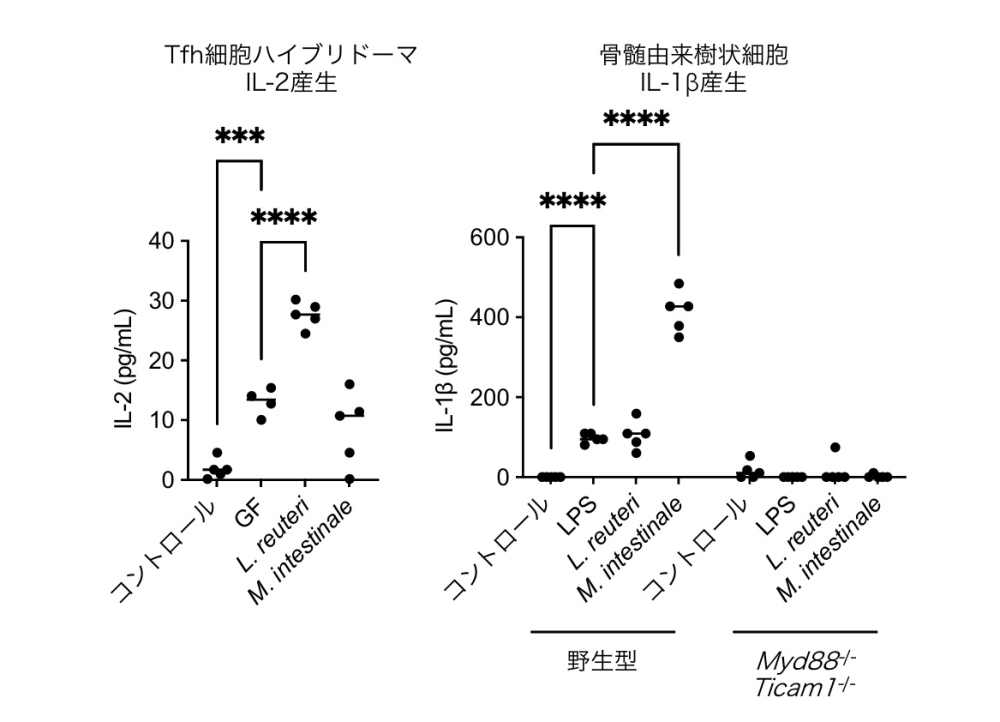
(左)パイエル板由来Tfh細胞ハイブリドーマを、GFマウス由来、 L. reuteri、またはM. intestinaleを定着させたマウス由来の小腸内容物でパルスした抗原提示細胞と共培養し、上清中のIL-2濃度をELISAで測定した。L. reuteri 刺激下でハイブリドーマからのIL-2産生が最も強く誘導され、L. reuteri 由来抗原がTfh細胞にcognateに認識されていることが示された。(右)野生型および Myd88−/−Ticam1−/− 二重欠損マウス (Toll様受容体シグナル伝達経路を欠失する)由来骨髄由来樹状細胞を、培地のみ(NC)、LPS (リポ多糖)、L. reuteri、M. intestinaleでそれぞれ刺激し、上清中の IL-1β産生量を測定した。野生型骨髄由来樹状細胞では M. intestinale刺激により著明な IL-1β産生が誘導されたが、Myd88−/−Ticam1−/− BMDC では完全に消失した。これはM. intestinale が MyD88/TRIF依存的な TLR シグナルを介して IL-1β産生を促進し、アジュバントとして機能することを示す。
(4)食事と菌のシナジーによる「食事+プロバイオティクス」戦略の実証
L. reuteriと M. intestinaleの保有量が低い飼育施設のマウスに対し、大豆食下で2菌を経口接種すると、パイエル板のTfh/IgA応答が回復しました。一方、精製飼料(AIN-93G)下では2菌の経口接種を行っても定着自体が成立せず、免疫応答も誘導されませんでした。この結果は、大豆由来の栄養環境が2菌種の定着土壌として必須であり、「食事」と「菌」が相乗的に作用して初めて腸管免疫が強化されることを示しています。すなわち、単なるプロバイオティクス投与ではなく、食事との組み合わせが鍵となることが明らかになりました(図4)。
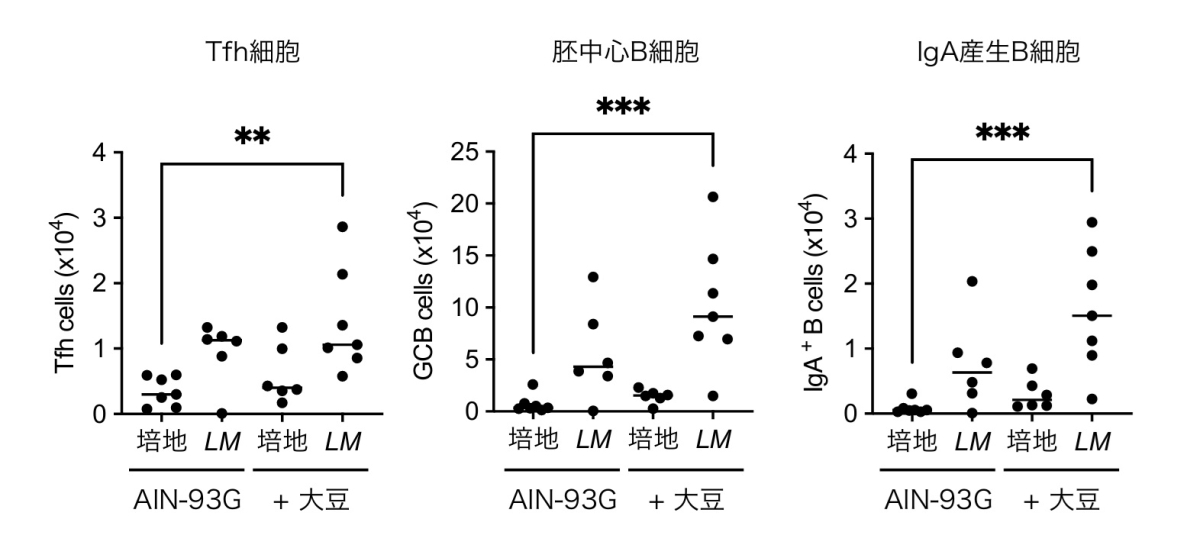
L. reuteriと M. intestinale の保有量が低い飼育施設由来マウスを用い、精製飼料(AIN-93G)または大豆添加AIN-93G飼料 (+大豆)の給餌下で、L. reuteriとM. intestinale混合菌液(LM)または細菌培養培地のみ(培地)またはを経口接種し、4週間後にパイエル板のTfh細胞、胚中心B細胞、IgA産生B細胞を解析した。AIN-93G下では L. reuteriとM. intestinale投与によってもTfh/IgA細胞は誘導されず、大豆食単独でも応答は不十分であり、大豆食と、 L. reuteriとM. intestinale経口接種を併用した群でのみ顕著な応答が認められた。これは食事(栄養環境)と腸内細菌の組み合わせが粘膜免疫の増強に必須であることを示す。
(5) 大豆食が誘導する多応答性IgAはサルモネラ菌感染を防御する
L. reuteri とM. intestinale ノトバイオートマウスから樹立したIgAハイブリドーマを解析したところ、複数のクローンが L. reuteriとM. intestinaleに加え、サルモネラ菌、大豆タンパク、LPS、フラジェリンなど多様な抗原に交差反応性を示す「多応答性IgA」であることが判明しました。IgA欠損マウスにこの多応答性IgA抗体を受動投与すると、サルモネラ菌感染に対する生存率が有意に延長されました(図5)。これにより、食事により誘導される多応答性IgAが、腸内細菌叢の恒常性維持と感染防御の両機能を併せ持つことが初めて実証されました(図6)。
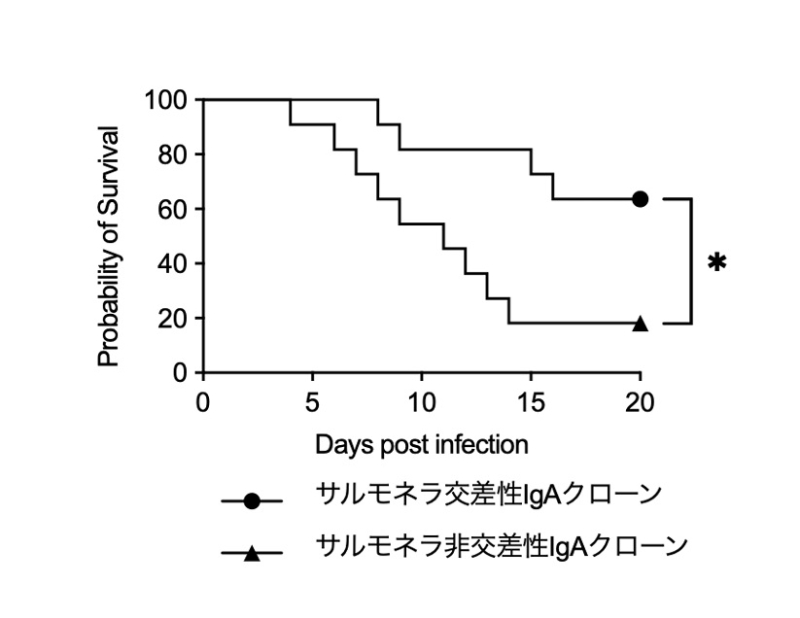
L. reuteriとM. intestinaleのノトバイオートマウスから樹立したIgAハイブリドーマから、Salmonella Typhimurium (サルモネラ)に交差反応性を示す多応答性IgAクローン(●)と、サルモネラに反応しない非交差性IgAクローン(▲)をそれぞれ選別した。各クローン由来モノクローナルIgAをIgA欠損マウスへ受動投与後、 S. Typhimuriumを経口感染させ、20日間の生存率を比較した。多応答性IgA投与群では非交差性IgA投与群と比較して生存率が有意に延長され、大豆食により誘導される多応答性IgAが共生細菌叢のみならず病原菌に対する感染防御も担うことが示された。
4. 今後の展望
・乳幼児期の栄養指導・予防医学への応用:離乳期の食事構成が長期的な腸管免疫の応答様式を規定するという知見は、離乳食・補完食の設計指針の科学的根拠となることが期待されます。
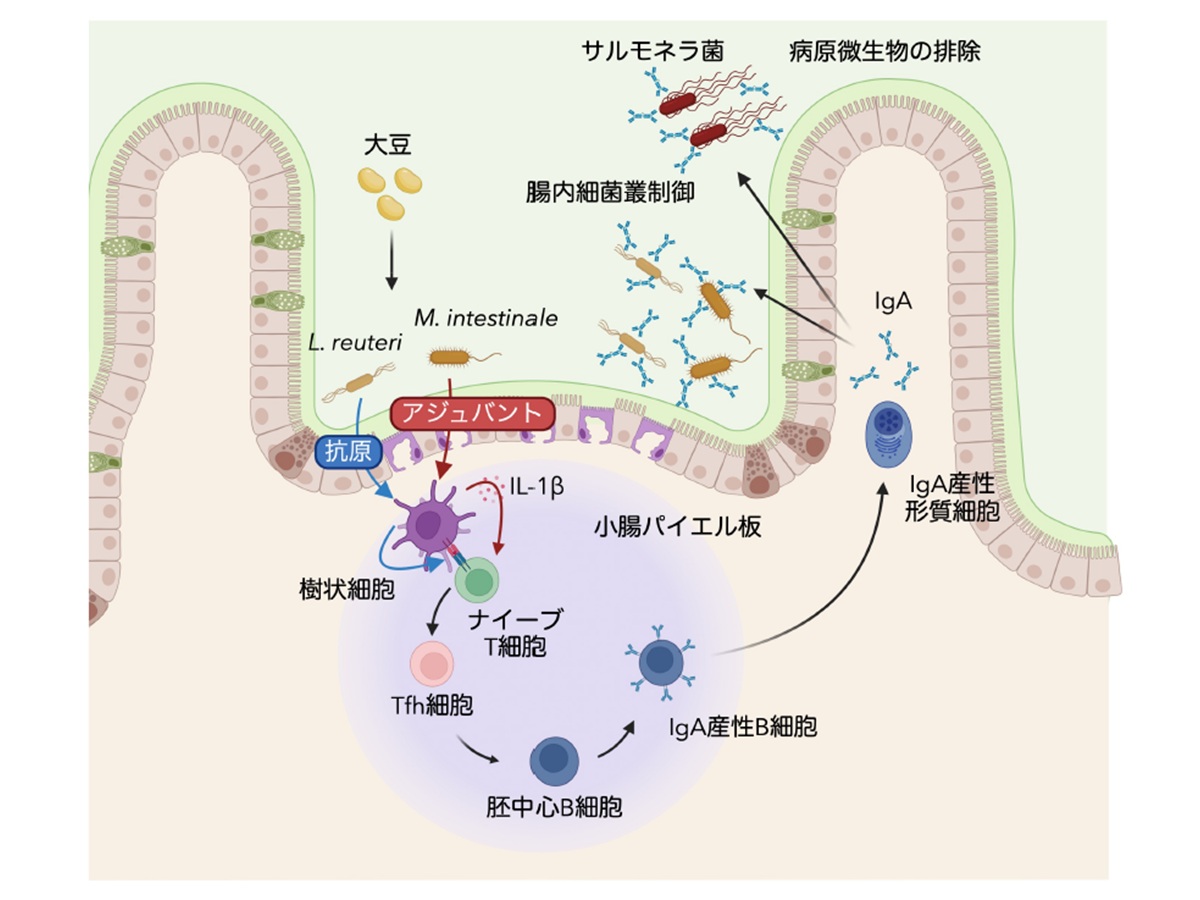
離乳期に大豆を摂取することで、小腸内で L. reuteri と M. intestinale が選択的に増殖・定着する。両菌はM細胞を介してパイエル板に取り込まれ、L. reuteri は Tfh 細胞に認識される抗原を提供し、M. intestinale は樹状細胞からの IL-1β 産生を介したアジュバントとして機能することで、Tfh細胞−胚中心B細胞−IgA産生形質細胞の連携を駆動する。誘導される分泌型IgAは共生細菌に加えS. Typhimuriumなどの病原菌にも結合する多応答性を示し、腸内細菌叢の恒常性維持と病原微生物の排除という二つの機能を併せ持つ。
5. 論文情報
【著者】Kisara Hattori-Muroi†, Hikari Maruta†, Daisuke Takahashi†*, Yusuke Kinashi, Kouya Hattori, Yuma Kabumoto, Yumiko Fujimura, Satoshi Tsukamoto, Koichiro Suzuki, Hiroyuki Oguchi, Yumi Ogihara, Yotaro Kodaira, Emi Hayashi, Kokona Takano, Seiga Komiyama, Naoki Morita, Hanako Naganawa-Asaoka, Yuki Oya, Yuka Saito, Wakana Ohashi, Shunsuke Kimura, Reiko Shinkura, Tsukasa Matsuda, Koji Hase* (†equal first authors, †*corresponding authors)
【掲載誌】Immunity
【DOI】 10.1016/j.immuni.2026.04.013
【URL】 https://www.cell.com/immunity/fulltext/S1074-7613(26)00175-5
6. 用語解説
※1パイエル板小腸の粘膜下に存在するリンパ組織で、腸管免疫応答の開始点として機能します。表面のM細胞が腸内細菌を取り込み、内部の免疫細胞に抗原を提示することで、IgA抗体の産生やT細胞の分化を誘導します。
※2濾胞性ヘルパーT(Tfh)細胞
リンパ組織の胚中心に局在するCD4陽性T細胞の一種で、B細胞の抗体産生を助ける役割を担います。転写因子Bcl-6とケモカイン受容体CXCR5の発現を特徴とし、高親和性抗体の産生に不可欠です。
※3多応答性IgA抗体
特定の1種類の抗原だけでなく、複数の異なる細菌種や分子構造に結合できるIgA抗体のことです。腸内細菌叢の広範な制御や、未知の病原菌に対する初期防御に寄与すると考えられています。
※4ノトバイオート実験
無菌環境で飼育したマウスに、特定の既知の細菌のみを人為的に定着させる実験です。個々の細菌が宿主の免疫系に与える影響を直接的に評価できる実験系として広く用いられています。
※5分泌型IgA抗体
腸管粘膜から管腔側に分泌される抗体で、腸内細菌叢の恒常性維持と病原体に対する初期防御に重要な役割を果たします。T細胞依存的経路で産生されるIgAは体細胞超変異を経て高親和性を獲得し、特異性と多応答性の両立を実現します。
※6 M細胞
M細胞は、パイエル板を覆う上皮層に存在する特殊な上皮細胞です。トランスサイトーシス機能が発達しており、管腔側から腸内細菌や食事由来成分などを取り込み、パイエル板内の抗原提示細胞へ受け渡す役割を担っています。