
| 発表のポイント |
|---|
|
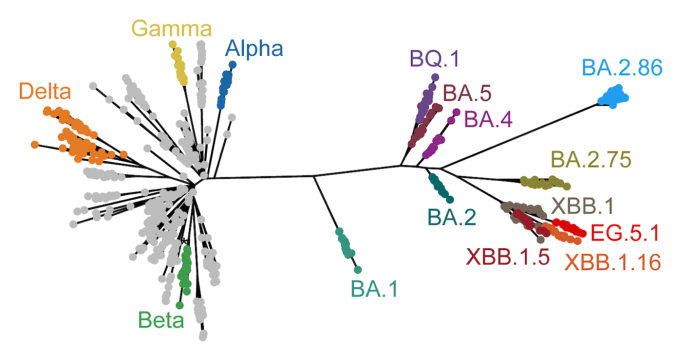
発表概要
東京大学医科学研究所システムウイルス学分野の佐藤佳教授が主宰する研究コンソーシアム「The Genotype to Phenotype Japan (G2P-Japan)」(注1)は、2023年8月ごろに出現したオミクロン(注2)BA.2.86株のウイルス学的特性を明らかにしました。オミクロンBA.2.86株は、それ以前に流行していたオミクロン亜株とは系統学的に大きく異なっており、オミクロンBA.2.86株のスパイクタンパク質(注3)には祖先株であるオミクロンBA.2株と比較して30種類以上の変異が蓄積しています。本研究では、オミクロンBA.2.86株は既存の流行株よりも有意に高い実効再生産数(注4)(流行拡大能力)を示すことを明らかにしました。さらに本研究では、オミクロンBA.2.86株についての詳細なウイルス学的実験を行いました。その結果、オミクロンBA.2.86株は、現在汎用されているレムデシビルやパキロビッド、ゾコーバなどの抗ウイルス薬のいずれに対しても高い感受性を有していました。感染受容体であるACE2(注5)結合能はEG.5.1株よりも高い一方で、合胞体形成活性(注6)、細胞での増殖能はEG.5.1株よりも低下していました。また、オミクロンBA.2.86株のハムスターモデルにおける病原性はオミクロンBA.2株、EG.5.1株に比べ、弱いことが明らかとなりました。
本研究成果は2024年1月26日、米国科学雑誌「Cell Host & Microbe」オンライン版で公開されました。
発表内容
ウイルス感染症の制御が難しい一因は、ウイルスが変異を獲得し進化することにあります。例えば、新型コロナウイルス(SARS-CoV-2)は流行の過程で著しく多様化し、「変異株」と呼ばれる様々な特性を持ったウイルスが出現してきました。SARS-CoV-2の研究を通し、ウイルスの進化と流行の原理を理解することができれば、新型コロナウイルス感染症(COVID-19)だけでなく、将来のパンデミックを含めた様々な感染症の制御に繋がる知見を得ることができると期待されます。2023年1月以降、遺伝子組換えにより出現したオミクロンXBB株およびその子孫株が、流行の中心を占めてきました。しかし、2023年8月にオミクロンBA.2.86株が出現して以降、流行の主流がオミクロンXBB株からオミクロンBA.2.86株へと移り変わりつつあります。オミクロンBA.2.86株は、オミクロンBA.2株の子孫株であるものの、オミクロンBA.2株と比較して、スパイクタンパク質に30ヶ所以上もの変異が認められます。既存の流行株と系統学的に大きく異なる変異株が突如出現するという観点から、オミクロンBA.2.86株の出現はオミクロン株の出現に匹敵する進化イベントであるとされています。オミクロンBA.2.86株は世界保健機関(WHO)より2023年8月に監視下にある変異株(variant under monitoring, VUM)に指定され、その後2023年11月に注目すべき変異株(variant of interest, VOI)に格上げされました。
本研究ではオミクロンBA.2.86株の流行拡大リスクを評価するため、まずウイルスゲノム疫学調査情報を基に、ヒト集団内におけるオミクロンBA.2.86株の実効再生産数を推定しました。その結果、オミクロンBA.2.86株の実効再生産数は、2023年11月当時の主流株であったオミクロンEG.5.1株(オミクロンXBB株の子孫株の1つ)の実効再生産数よりも高いことが分かりました(図1)。これは、オミクロンBA.2.86株が今後流行を拡大していき、主流行株のひとつになる可能性が高いことを示しています。
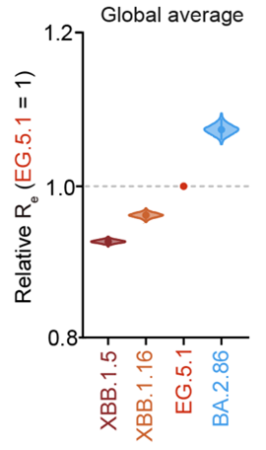
ウイルスゲノム疫学調査の結果から数理モデルを用いて各変異株の実効再生産数(伝播力の指標)を推定した。縦軸はオミクロンEG.5.1株の値を基準としたときの実効再生産数を示す。値が大きいほどウイルスの伝播力が高いことを示す。
本研究ではさらに、オミクロンBA.2.86株についての詳細なウイルス性状解析実験を行いました。オミクロンBA.2.86株はEG.5.1株およびBA.2.86株の祖先株であるBA.2株よりもACE2結合能が向上していた一方で(図2a)、オミクロンBA.2.86株の合胞体形成活性(図2b)および培養細胞での増殖能(図2c)はEG.5.1株よりも低いことがわかりました。また、生体での呼吸器系の環境を模倣する気道チップを用いた実験の結果、オミクロンBA.2.86株は EG.5.1株よりも呼吸器上皮―血管内皮バリア破壊能が低いことがわかりました(図2d)。さらに、オミクロンBA.2.86株に対して、現在汎用されているレムデシビルやパキロビッド、ゾコーバなどの抗ウイルス薬はいずれも効果的であることを確認しました(図2e)。
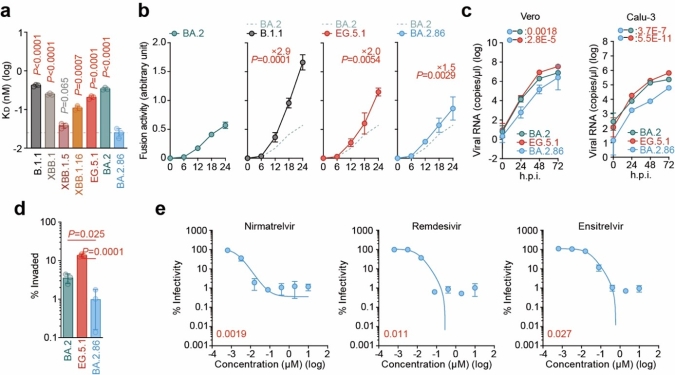
a)オミクロンBA.2.86株のACE2結合活性。KD値が低いほど、ACE2に対する結合活性が高いことを示す。b) オミクロンBA.2.86株の合胞体形成活性。c) オミクロンBA.2.86株のVero細胞およびCalu-3細胞での増殖試験。d)気道チップを用いたオミクロンBA.2.86株の気道上皮―内皮バリア破壊能の評価。e)オミクロンBA.2.86株に対するパキロビッド(Nirmatrelvir)やレムデシビル(Remdesivir)、ゾコーバ(Ensitrelvir)の抗ウイルス効果を検証。
最後に、ハムスターモデルを用いた感染実験により、オミクロンBA.2.86株の個体内感染動態と病原性を評価しました。その結果、オミクロンBA.2.86株のハムスターの肺における増殖力は、祖先型であるオミクロンBA.2株およびEG.5.1株よりも低下しており、また病原性も低いことが明らかとなりました(図3)。
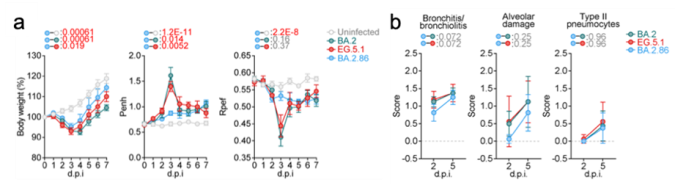
a) ウイルスに感染したハムスターの体重、肺機能の推移。
b)ハムスター肺の組織病理学的スコア。値が高いほど、炎症や肺障害の程度が強いことを示す。
本研究で解析したオミクロンBA.2.86株は、その後さらに多様化し、進化しました。2024年1月現在、オミクロンBA.2.86株の子孫株であるオミクロンJN.1株が世界的な感染拡大を引き起こしており、問題となっています。今後もオミクロンBA.2.86株およびその子孫株の流行、性質、進化を継続的に監視することが重要です。
G2P-Japanでは、SARS-CoV-2の進化・流行動態を司る原理の解明に関する研究、および、出現が続くさまざまな変異株について、ウイルス学的な特性の解析や、中和抗体や治療薬への感受性の評価、病原性についての研究に取り組んでいます。G2P-Japanコンソーシアムでは、今後も、SARS-CoV-2の変異(Genotype)の早期捕捉と、その変異がヒトの免疫やウイルスの病原性・複製に与える影響(Phenotype)を明らかにするための研究を推進します。
発表者
東京大学医科学研究所 感染・免疫部門 システムウイルス学分野佐藤 佳(教授, 一般社団法人G2P-Japan 代表理事)
伊東 潤平(准教授)
瓜生 慧也(大学院生)
Jarel Elgin M. Tolentino(大学院生)
北海道大学 大学院医学研究院
田中 伸哉(教授)
福原 崇介(教授, 一般社団法人G2P-Japan 理事)
田村 友和(講師)
小田 義崇(助教)
北海道大学 人獣共通感染症国際共同研究所
松野 啓太(准教授)
直 亨則(特任講師)
水間 奎太(博士研究員)
児島 一州(JSPS特別研究員)
京都大学 iPS細胞研究所(CiRA)
高山 和雄(講師)
出口 清香(大学院生)
広島大学 大学院医系科学研究科
入江 崇(准教授)
ヒトレトロウイルス学共同研究センター 熊本大学キャンパス
池田 輝政(准教授)
Hesham Nasser(特任助教)
宮崎大学 農学部獣医学科
齊藤 暁(准教授)
田中 友理(大学生)
論文情報
〈雑誌〉Cell Host & Microbe〈題名〉Virological characteristics of the SARS-CoV-2 BA.2.86 variant
〈著者〉田村友和#, 水間奎太#, Hesham Nasser#, 出口清香#, Miguel Padilla-Blanco#, 小田義崇#, 瓜生慧也#, Jarel E.M. Tolentino#, 辻野修平, 鈴木理滋, 児島一州, 直亨則, 清水凌, 王磊, 津田真寿美, Michael Jonathan, 小杉優介, Ziyi Guo, Alfredo A. Hinay Jr., Olivia Putri, Yoonjin Kim, 田中友理, 浅倉弘幸, 長島真美, 貞升健志, 吉村和久, The Genotype to Phenotype Japan (G2P-Japan) Consortium, 齊藤暁, 伊東潤平, 入江崇, 田中伸哉*, Jiri Zahradnik*, 池田輝政*, 高山和雄*, 松野啓太*, 福原崇介*, 佐藤佳*
(#Equal contribution; *Corresponding author)
〈DOI〉10.1016/j.chom.2024.01.001
〈URL〉https://www.sciencedirect.com/science/article/pii/S1931312824000052
研究助成
本研究は、佐藤 佳教授に対する日本医療研究開発機構(AMED)「新興・再興感染症に対する革新的医薬品等開発推進研究事業(JP22fk0108146, JP21fk0108494, JP21fk0108425, JP21fk0108432)」、AMED 先進的研究開発戦略センター(SCARDA)「ワクチン開発のための世界トップレベル研究開発拠点の形成事業(UTOPIA, JP223fa627001)、AMED SCARDA「ワクチン・新規モダリティ研究開発事業(JP223fa727002)」、科学技術振興機構(JST) CREST(JPMJCR20H4)、伊東 潤平助教に対するJSTさきがけ(JPMJPR22R1)などの支援の下で実施されました。用語解説
(注1)研究コンソーシアム「The Genotype to Phenotype Japan (G2P-Japan)」東京大学医科学研究所 システムウイルス学分野の佐藤佳教授が主宰する研究コンソーシアム。日本国内の様々な専門性を持つ若手研究者が参画し、多角的アプローチからウイルスの性質の解明に取り組んでいる。現在では、イギリスを中心とした諸外国の研究チーム・コンソーシアムとの国際連携も進めている。
(注2)オミクロン株(B.1.1.529, BA系統)
新型コロナウイルスの流行拡大によって出現した、顕著な変異を有する「懸念すべき変異株(VOC:variant of concern)」のひとつ。オミクロンBA.1株、オミクロンBA.5株、オミクロンBQ.1.1株、オミクロンXBB株などが含まれる。現在、日本を含めた世界各国で大流行している変異株である。
(注3)スパイクタンパク質
新型コロナウイルスが細胞に感染する際に、新型コロナウイルスが細胞に結合するためのタンパク質。現在使用されているワクチンの標的となっている。
(注4)実効再生産数
特定の状況下において、1人の感染者が生み出す二次感染者数の平均。ここでは、変異株間の流行拡大能力の比較の指標として用いている。
(注5)ACE2
Angiotensin-Converting Enzyme 2(アンジオテンシン変換酵素2)の略称で、新型コロナウイルスが細胞に感染する際に受容体として機能する。
(注6)合胞体形成活性
合胞体とは、新型コロナウイルスに感染した細胞が、スパイクタンパク質を細胞表面に発現し、周囲の細胞と融合することによって形成される大きな細胞塊のこと。合胞体形成活性とは、新型コロナウイルスのスパイクタンパク質を介して、合胞体を形成する能力のこと。
問合せ先
〈研究に関する問合せ〉東京大学医科学研究所 感染・免疫部門 システムウイルス学分野
教授 佐藤 佳(さとう けい)
https://www.ims.u-tokyo.ac.jp/imsut/jp/lab/ggclink/section04.html
広島大学大学院医系科学研究科 ウイルス学
准教授 入江 崇(いりえ たかし)
https://seeds.office.hiroshima-u.ac.jp/profile/ja.8749775bdccf6f20520e17560c007669.html
〈報道に関する問合せ〉
東京大学医科学研究所 プロジェクトコーディネーター室(広報)
https://www.ims.u-tokyo.ac.jp/
北海道大学 社会共創部広報課
https://www.hokudai.ac.jp/
京都大学iPS細胞研究所(CiRA) 国際広報室
https://www.cira.kyoto-u.ac.jp/
広島大学 広報室
https://www.hiroshima-u.ac.jp/
熊本大学 総務部総務課広報戦略室
https://www.kumamoto-u.ac.jp/
宮崎大学 企画総務部 総務広報課
https://www.miyazaki-u.ac.jp/