
| 発表のポイント |
|---|
|
発表内容
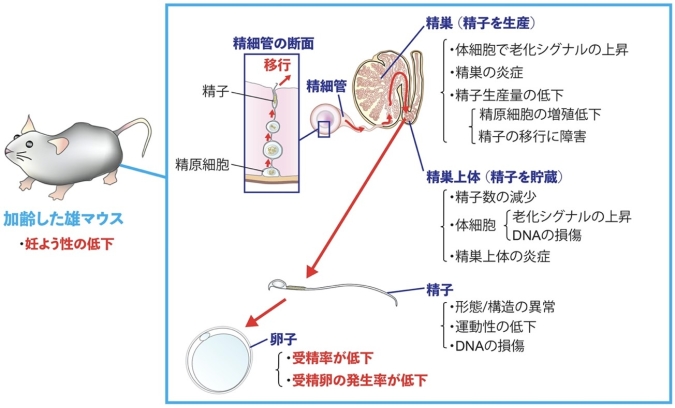
東京大学大学院農学生命科学研究科の遠藤墾助教は、大阪大学微生物病研究所の伊川正人教授(兼: 東京大学医科学研究所 特任教授)らによる研究グループと共同で、精子をつくる精巣と精子を貯蔵する精巣上体がそれぞれ老化し、精子や受精卵の発育に悪影響を及ぼすことを発見しました。〈研究の背景〉
近年の晩婚化を背景に、ヒトでは男性の加齢による精子や妊よう性(注1)への影響が注目されています。また、牛や豚を含む畜産動物でも、加齢した雄の繁殖障害として、精子の数や質の低下が知られ、雌の受胎率低下のリスクとして認識されています。 しかしヒトや畜産動物では、非侵襲性の精液検査や微量の精巣組織採取(バイオプシー)による集積的な症例報告が主であり、詳細な解析が困難でした。またヒトでは、生活習慣や環境感受性の個人差も大きく、加齢による精子や妊よう性の変化が、生活習慣・環境因子の長期的な影響によるものか、生物学的な老化現象によるものかは不明瞭でした。
〈研究の内容〉
本研究グループは、ヒトを含め雄性哺乳類の加齢における、妊よう性や精子機能の変化とそれぞれの相関性について、遺伝的・環境的に統制された近交系(注2)の雄マウスを用いて網羅的に解析し、これらの変化の特定とその原因究明を試みました。
雄マウスを若齢期から老齢期まで、若い雌と交配させ続けたところ、雄の加齢とともに雌が出産する仔の数が徐々に減少しており、この減少(妊よう性の低下)に雄の交尾頻度や雄性ホルモンであるテストステロン(注3)量は関連がありませんでした。そこで次に、精子を生産する器官である精巣と、精巣でつくられた精子を受精までのあいだ貯蔵する器官である精巣上体に着目したところ、精巣と精巣上体それぞれの体細胞で老化関連β-ガラクトシダーゼ活性(注4)が上昇し、老化細胞による両器官の炎症がみられました。したがって、精巣や精巣上体の老化による機能低下が、妊よう性の低下の原因であると考えられました。
加齢した精巣を構成する精細管(注5)の内部では、精子の元となる精原細胞(注6)の増殖が低下することで、生産される精子数が減少していました。また、つくられた精子を精細管の内腔に放出して精巣上体に移行させる機能が低下し、精巣上体の精子数の減少につながることを発見しました。加齢した精巣上体では、貯蔵された精子の形態や微細構造に異常が蓄積していき、このため精子の運動性も低下し、受精率(注7)低下の原因となっていました。
続いて、加齢精子と受精できた卵子に着目し、その後の発生(発育)を観察したところ、発生率(注8)も低下していましたが、精子の受精率低下と受精卵の発生率低下は相関しないことを発見しました。これにより、加齢した精巣上体は精巣よりもDNA損傷を受けやすい内部環境となっており、その環境下で貯蔵された精子にDNA損傷が蓄積し、発生率低下の原因となることがわかりました。
〈今後の展望〉
以上の結果から、精巣・精巣上体の老化が精子の受精率低下と受精卵の発生率低下を独立して引き起こし、妊よう性の低下に至ることを突き止めました。本成果は、雄の加齢と妊よう性、および精子の質・量の低下との相関性、さらにその実態を生物学的に解明した重要な発見であるといえます。
加齢による妊よう性低下は、男性(雄)の生殖器官の老化にも起因することがわかりました。今後は、加齢男性や雄の畜産動物にみられる妊よう性低下の原因理解とともに、新たな治療技術や予防技術の確立につながることが期待されます。
発表者・研究者等情報
東京大学
大学院農学生命科学研究科
遠藤 墾 助教 兼:東京医科歯科大学実験動物センター 非常勤講師
医科学研究所
小沢 学 准教授
石川 祐 研究当時:東京大学特別研究員(日本学術振興会特別研究員PD)
現:横浜市立大学医学部 助教
大阪大学 微生物病研究所
伊川 正人 教授 兼:東京大学医科学研究所 特任教授
原 英二 教授
宮田 治彦 准教授
河本 新平 准教授
奥崎 大介 特任准教授
元岡 大祐 講師
江森 千紘 助教
嶋田 圭祐 助教
松村 貴史 研究当時:博士課程
小林 清訓 研究当時:博士課程
島田 健太郎 研究当時:修士課程
小谷 まよ 研究当時:修士課程
論文情報
雑誌名:Communications Biology
題 名:Multiple ageing effects on testicular/epididymal germ cells lead to decreased male fertility in mice
著者名:Tsutomu Endo*, Kiyonori Kobayashi, Takafumi Matsumura, Chihiro Emori, Manabu Ozawa, Shimpei Kawamoto, Daisuke Okuzaki, Keisuke Shimada, Haruhiko Miyata, Kentaro Shimada, Mayo Kodani, Yu Ishikawa-Yamauchi, Daisuke Motooka, Eiji Hara, Masahito Ikawa* (*責任著者)
DOI:10.1038/s42003-023-05685-2
URL:https://doi.org/10.1038/s42003-023-05685-2
題 名:Multiple ageing effects on testicular/epididymal germ cells lead to decreased male fertility in mice
著者名:Tsutomu Endo*, Kiyonori Kobayashi, Takafumi Matsumura, Chihiro Emori, Manabu Ozawa, Shimpei Kawamoto, Daisuke Okuzaki, Keisuke Shimada, Haruhiko Miyata, Kentaro Shimada, Mayo Kodani, Yu Ishikawa-Yamauchi, Daisuke Motooka, Eiji Hara, Masahito Ikawa* (*責任著者)
DOI:10.1038/s42003-023-05685-2
URL:https://doi.org/10.1038/s42003-023-05685-2
研究助成
本研究は、科研費「新学術領域研究 (課題番号:19H05750)」、科研費「基盤研究(S)(課題番号:21H05033)」、科研費「基盤研究(C)(課題番号:19K06439,23K05587)」、国立研究開発法人日本医療研究開発機構(AMED)「老化メカニズムの解明・制御プロジェクト(課題番号:21gm5010001)」の支援により実施されました。
用語解説
(注1)妊よう性
妊娠し、子供を授かるために必要な能力。女性(雌)と男性(雄)のどちらにも関係する。
(注2)近交系
遺伝的に均一な実験動物の系統のこと。個体差による実験成績のばらつきが少なくするために用いられる。
(注3)テストステロン
主要な男性(雄性)ホルモンであり、動物では交尾行動に影響を与えることが知られる。臨床現場では、総テストステロン量と遊離テストステロン量が測定可能だが、加齢によりどちらの量も低下することが報告されている(SM Harmanら, J Clin Endocrinol Metab, 6(2):724-31, 2001; DOI:10.1210/jcem.86.2.7219)。
(注4)老化関連β-ガラクトシダーゼ活性
老化に関連して細胞内のβ-ガラクトシダーゼ活性が上昇するため、細胞老化を検出するためのバイオマーカーとして広く用いられる。
(注5)精細管
精巣を構成する、精子を生産する場となる管。管内部の上皮組織でつくられた精子は、管内腔に放出され、精巣上体へと移行する。
(注6)精原細胞
精子の元となる未分化な生殖細胞。精原細胞は増殖して数を増やしつつ、その一部が精子となることで、精巣が大量の精子を継続して生産することが可能となる。
(注7)受精率
排卵された正常な卵子の総数に対して、精子と受精することができた卵子の割合を示す。
(注8)発生率
受精した卵子は受精卵(または胚)と呼ばれ、この受精卵が新しい生命として発生(発育)する。発生率は、受精卵の総数に対して、着床前の段階(胚盤胞)まで正常に発生できた割合を示す。着床が完了すると、妊娠が成立する。
妊娠し、子供を授かるために必要な能力。女性(雌)と男性(雄)のどちらにも関係する。
(注2)近交系
遺伝的に均一な実験動物の系統のこと。個体差による実験成績のばらつきが少なくするために用いられる。
(注3)テストステロン
主要な男性(雄性)ホルモンであり、動物では交尾行動に影響を与えることが知られる。臨床現場では、総テストステロン量と遊離テストステロン量が測定可能だが、加齢によりどちらの量も低下することが報告されている(SM Harmanら, J Clin Endocrinol Metab, 6(2):724-31, 2001; DOI:10.1210/jcem.86.2.7219)。
(注4)老化関連β-ガラクトシダーゼ活性
老化に関連して細胞内のβ-ガラクトシダーゼ活性が上昇するため、細胞老化を検出するためのバイオマーカーとして広く用いられる。
(注5)精細管
精巣を構成する、精子を生産する場となる管。管内部の上皮組織でつくられた精子は、管内腔に放出され、精巣上体へと移行する。
(注6)精原細胞
精子の元となる未分化な生殖細胞。精原細胞は増殖して数を増やしつつ、その一部が精子となることで、精巣が大量の精子を継続して生産することが可能となる。
(注7)受精率
排卵された正常な卵子の総数に対して、精子と受精することができた卵子の割合を示す。
(注8)発生率
受精した卵子は受精卵(または胚)と呼ばれ、この受精卵が新しい生命として発生(発育)する。発生率は、受精卵の総数に対して、着床前の段階(胚盤胞)まで正常に発生できた割合を示す。着床が完了すると、妊娠が成立する。
問合せ先
(研究内容については発表者にお問合せください)
東京大学 大学院農学生命科学研究科 応用動物科学専攻
助教 遠藤 墾(えんどう つとむ)
東京大学 大学院農学生命科学研究科 広報室
東京大学 医科学研究所 プロジェクトコーディネーター室(広報)