
| 発表のポイント |
|---|
|
発表概要
東京大学医科学研究所システムウイルス学分野の佐藤佳教授が主宰する研究コンソーシアム「The Genotype to Phenotype Japan (G2P-Japan)」(注1)は、オミクロン株(注2)の進化過程において適応度(流行拡大能力)(注3)の上昇に寄与した変異を同定しました。進化系統解析により、様々なオミクロン亜株が、スパイクタンパク質(注4)の重要な5箇所のアミノ酸残基において変異を収斂(注5)的に獲得していることを明らかにしました。次に、流行モデリング解析により、前述の5箇所の変異(収斂変異)の獲得が、ウイルスの適応度を上昇させることを示しました。さらに、i) 収斂変異を多く獲得した変異株ほど高い適応度を示すこと、ii) 5箇所の収斂変異の獲得により、オミクロン株の進化過程における適応度上昇の大部分が説明できることを示しました。
これらの結果は、一見複雑に見えるオミクロン株の進化パターンが、多くの収斂変異を獲得するほど適応度が上昇するという、単純な法則で説明可能であることを示しています。
さらに本研究では、5箇所全ての収斂変異を獲得し、高い適応度を示すオミクロン「BQ.1.1株」について、詳細なウイルス学的実験を行いました。その結果、オミクロンBQ.1.1株は収斂変異を獲得したことで、祖先株であるオミクロンBA.5株よりも高いACE2(注6)結合能、感染性、そして液性免疫(注7)に対する逃避能を獲得したことが明らかとなりました。一方で、オミクロンBQ.1.1株の実験動物モデルにおける病原性はオミクロンBA.5株と同程度であることが明らかとなりました。本研究成果は2023年5月11日、英国科学雑誌「Nature Communications」オンライン版に掲載されました。
発表内容
ウイルス感染症の制御が難しい一因は、ウイルスが変異を獲得し進化することにあります。例えば、新型コロナウイルス(SARS-CoV-2)は流行の過程で著しく多様化し、「変異株」と呼ばれる様々な特性を持ったウイルスが出現してきました。現在でも、ワクチン接種や自然感染により獲得された液性免疫に対する高い逃避能を持った変異株が次々と出現してきており、医学・公衆衛生上の問題となっています。新型コロナウイルスの研究を通し、ウイルスの進化と流行の原理を理解することができれば、新型コロナウイルス感染症だけでなく、将来のパンデミックを含めた様々な感染症の制御に繋がる知見を得ることができると期待されます。2021年末に出現したオミクロン株(B.1.1.529, BA系統)は、流行の過程で著しく多様化し、様々なオミクロン亜株(オミクロンBA.2株、BA.5株、BA.2.75株、BQ.1.1株など)が出現してきました(図1)。これらのオミクロン亜株は、より遅くに出現したものほど高い適応度(流行拡大能力)を有しており、オミクロン株の適応度が進化の過程で徐々に上昇しつつあることが指摘されています。しかし、このような適応度の上昇が、どの様な進化的メカニズムにより引き起こされるのか、明らかでありませんでした。
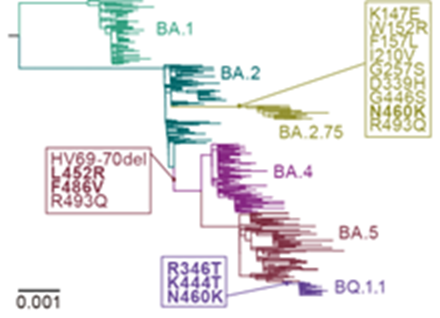
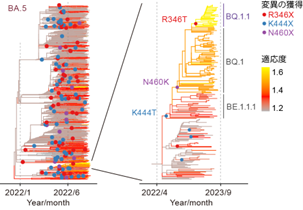
図はオミクロン亜株の系統樹を表す。オミクロンBA.1株が出現した後、様々なオミクロン亜株が継続的に出現し、流行を引き起こしてきた。オミクロンBA.4株とBA.5株の共通祖先、オミクロンBA.2.75株、およびオミクロンBQ.1.1株において新たに獲得されたスパイクタンパク質の変異を表示している。太字は収斂変異の獲得を示す。
重要なことに、様々なオミクロン亜株が、スパイクタンパク質の受容体結合ドメイン(注8)に存在する5箇所のアミノ酸において共通の変異を持つことが知られています(R346X, K444X, L452X, N460X, F486X; Xは任意のアミノ酸)(図2)。この現象は、これらの変異がオミクロン株の進化・多様化の過程で繰り返し(すなわち収斂的に)獲得されてきたことを示唆しています。しかし、これらの変異(以下、収斂変異と呼称)がどの程度高頻度に獲得されてきたか、そしてこれらの変異がなぜ高頻度に獲得されてきたのか、については不明でした。

本稿では、スパイクタンパク質配列に存在する346番目のアルギニン(R; R346)、444番目のリジン(K; K444)、452番目のロイシン(L; L452)、460番目のアスパラギン(N; N460)、486番目のフェニルアラニン(F; F486)における変異を収斂変異と呼称する。これらの収斂変異は、様々なオミクロン亜株において独立に複数回(すなわち収斂的に)獲得されている。例;オミクロンBA.2.75株とオミクロンBQ.1.1株におけるN460K変異。
そこで本研究では、オミクロン株のスパイクタンパク質における収斂進化を解析することで、オミクロン株の進化パターンの解明を試みました。まず、オミクロン株を対象とした大規模な系統解析を実施しました(図3)。
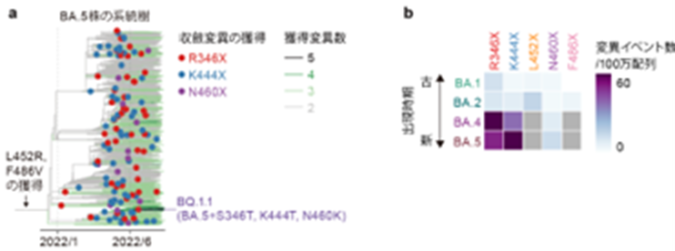
a) オミクロンBA.5株における収斂変異の獲得イベントの推定。オミクロンBA.5株の系統樹において、各収斂変異が獲得された枝を推定した。
b) オミクロンBA.1株、BA.2株、BA.4株、BA.5株における収斂変異の獲得頻度を表すヒートマップ。灰色は共通祖先の段階で既に獲得されていることを示している。オミクロン株の全ての系統において、収斂変異は繰り返し獲得されているが、より後に出現した系統ほど収斂変異の獲得頻度が高い傾向にある。
そして、各変異の獲得が系統樹上で何回起こったかを推定することで、オミクロン株の進化過程において収斂変異がどの程度の頻度で獲得されたか解析しました。その結果、これらの収斂変異はオミクロン株の比較的若い系統(オミクロンBA.4株およびBA.5株)において特に高頻度に獲得されてきたことを明らかにしました。
次に、前述の5つの収斂変異がオミクロン株の流行拡大能力(適応度)に与える影響を解析しました(図4)。そのために、変異株の流行動態に基づき、ウイルスの持つ各変異が適応度に与える影響を推定する数理モデルを開発しました。このモデルを用いると、各変異の適応度に与える影響(変異の獲得が適応度をどの程度増減させるか)だけでなく、ウイルスの持つ変異パターンからそのウイルスの適応度を予測することができます。
このモデルを用いた解析の結果、1) 収斂変異の獲得はオミクロン株の適応度を大きく上昇させること、2) 多くの種類の収斂変異を持つオミクロン亜株ほど高い適応度を示すこと、3) 特に、オミクロンBA.5株の子孫であり、全ての収斂変異を獲得したオミクロンBQ.1.1株が、解析したオミクロン亜株の中で最も高い適応度を示すことを明らかにしました(図4)。さらに、スパイクタンパク質における5箇所の収斂変異に基づき、オミクロン株の進化過程で見られる適応度の上昇の約80%が説明できることを示しました(図4)。
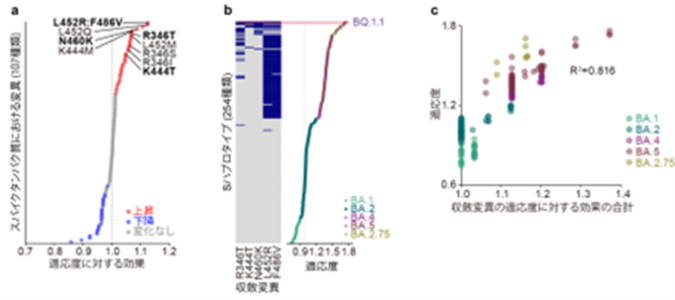
スパイクタンパク質における変異のパターンに基づき、オミクロン株のゲノム配列を254種類のグループ(スパイクタンパク質(S)ハプロタイプ)に分類した。その後、Sハプロタイプの流行動態から、i) 各変異の有無が適応度に与える効果と、ii) 各Sハプロタイプの適応度を同時に推定する数理モデルを開発し、これらの値を推定した。
a) 各変異がオミクロン株の適応度に与える効果の推定。収斂変異については変異名を表示している。収斂変異はスパイクタンパク質における変異の中で、トップクラスに適応度を上昇させる効果が高い。
b) 各Sハプロタイプの適応度の推定。点の色は各SハプロタイプのPANGO系統を表す。左側のヒートマップは、各Sハプロタイプにおける5種類の収斂変異の有無を表す。
c) オミクロン株の適応度変化における収斂変異の重要性。縦軸は適応度を、横軸は収斂変異の適応度に対する効果の合計を表す。オミクロン株の進化過程において見られる適応度の変化の大部分(約80%)は、収斂変異の獲得により説明できることを示す。
本研究ではさらに、収斂変異を全て獲得し、解析した中で最も高い適応度を示すオミクロンBQ.1.1株について、詳細なウイルス性状解析実験を行いました(図5)。オミクロンBQ.1.1株は、オミクロンBA.5株(L452RとF486Vの2つの収斂変異を持つ)がさらに3つの収斂変異(R346T、K444T、N460K)を獲得したことにより生じた変異株です(図1)。中和試験(注9)の結果、オミクロンBQ.1.1株は祖先型であるオミクロンBA.5株と比較して、ワクチン接種やブレークスルー感染(注10)により獲得された中和抗体に対し高い逃避能を示すことが明らかとなりました(図5)。
また、オミクロンBQ.1.1株のスパイクタンパク質は、オミクロンBA.5株と比較して感染受容体であるACE2への結合力が高いことが明らかとなりました(図5)。さらに、シュードウイルス(注11)を用いた感染実験において、オミクロンBQ.1.1株のスパイクタンパク質は、オミクロンBA.5株よりも高い感染力を示すことが明らかとなりました(図5)。また、オミクロンBQ.1.1株はオミクロンBA.5株よりも高い合胞体形成活性(注12)を示すことが明らかになりました。
以上の結果から、オミクロンBQ.1.1株が祖先型であるオミクロンBA.5株よりも高い適応度を示すのは、3つの収斂変異を追加で獲得することにより、液性免疫に対する逃避能、ACE2結合能、そして感染性を上昇させたことが原因であることが示唆されました。
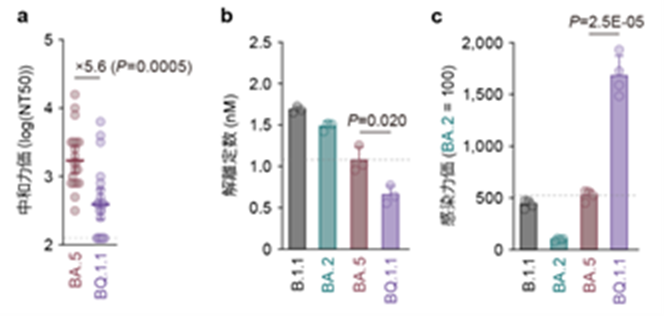
a) オミクロンBA.5株のブレークスルー感染者血清を用いたウイルス中和試験。中和力価の低いほどウイルスの中和抗体に対する逃避能が高いことを意味する。
b) 感染受容体であるACE2とスパイクタンパク質の結合強度。解離定数が低いほどACE2 に対する結合強度が高いことを意味する。
c) シュードウイルスを用いた感染性の評価。
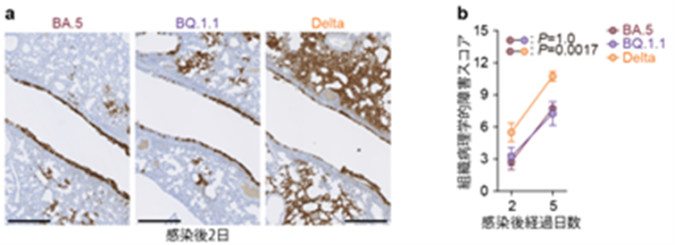
a) 感染後2日目のハムスター肺におけるウイルスの増殖。ウイルスが感染している細胞(Nタンパク質陽性細胞)は、茶色に免疫染色されている。
b)ハムスター肺の組織病理学的障害スコア。値が高いほど組織障害の程度が強いことを示す。
発表者
東京大学医科学研究所 システムウイルス学分野佐藤 佳(教授)
伊東 潤平(助教)
瓜生 慧也(大学院生)
北海道大学 大学院医学研究院
田中 伸哉(教授)
福原 崇介(教授)
鈴木 理滋(助教)
北海道大学 創成研究機構 化学反応創成研究拠点(ICReDO)
王 磊(特任助教)
北海道大学 人獣共通感染症国際共同研究所
松野 啓太(准教授)
板倉 友香里(大学院生)
京都大学 医生物学研究所
橋口 隆生(教授)
木村 香菜子(助教)
京都大学 iPS細胞研究所
高山 和雄(講師)
出口 清香(大学院生)
熊本大学 ヒトレトロウイルス学共同研究センター
池田 輝政(准教授)
宮崎大学 農学部獣医学科
齊藤 暁(准教授)
HiLung株式会社
山本 祐樹(代表取締役)
永元 哲治(取締役)
論文情報
研究助成
本研究は、佐藤佳教授らに対する日本医療研究開発機構(AMED)「新興・再興感染症に対する革新的医薬品等開発推進研究事業(JP22fk0108146, JP21fk0108494, JP21fk0108425, JP21fk0108432)」、AMED 先進的研究開発戦略センター(SCARDA)「ワクチン開発のための世界トップレベル研究開発拠点の形成事業(UTOPIA, JP223fa627001)、AMED SCARDA「ワクチン・新規モダリティ研究開発事業(JP223fa727002)」、科学技術振興機構(JST) CREST(JPMJCR20H4)、および伊東潤平助教に対する科学技術振興機構 さきがけ(JPMJPR22R1)などの支援の下で実施されました。用語解説
(注1)研究コンソーシアム「The Genotype to Phenotype Japan (G2P-Japan)」東京大学医科学研究所 システムウイルス学分野の佐藤教授が主宰する研究コンソーシアム。日本国内の様々な専門性を持つ若手研究者が参画し、多角的アプローチからウイルスの性質の解明に取り組んでいる。現在では、イギリスを中心とした諸外国の研究チーム・コンソーシアムとの国際連携も進めている。
(注2)オミクロン株
新型コロナウイルスの流行拡大によって出現した、顕著な変異を有する「懸念すべき変異株(VOC:variant of concern)」のひとつ。オミクロンBA.1株、オミクロンBA.2株、オミクロンBA.5株、オミクロンBQ.1.1株などが含まれる。現在、日本を含めた世界各国で大流行しており、パンデミックの主たる原因となる変異株となっている。
(注3)適応度(流行拡大能力 )
生物がどれだけ多くの子孫を次世代に残すことができるかを示す尺度。ウイルスの場合、特定の条件下において1人のウイルス感染者が平均して何人の二次感染者を生み出せるかを表す尺度。実効再生産数と同義。本研究では変異株の流行拡大能力を比較する目的で使用している。
(注4)スパイクタンパク質
新型コロナウイルスが細胞に感染する際に、新型コロナウイルスが細胞に結合するためのタンパク質。現在使用されているワクチンの標的となっている。
(注5)収斂
系統の異なる生物が、類似の形質を独立に獲得すること。
(注6)ACE2
Angiotensin-Converting Enzyme 2(アンジオテンシン変換酵素2)の略称で、新型コロナウイルスが細胞に感染する際に受容体として機能する。
(注7)液性免疫
B細胞が主体となって、抗体を作ることで異物に対抗する免疫。
(注8)受容体結合ドメイン
スパイクタンパク質のうち、感染受容体であるACE2との結合に重要な領域。
(注9)中和試験
抗体や抗体を含む血清がウイルスを不活化(中和)する能力を定量するための実験。ウイルスを中和する能力のある抗体は中和抗体と呼ばれる。新型コロナウイルスの場合、中和抗体はスパイクタンパク質に結合することでスパイクタンパク質と感染受容体であるACE2との相互作用を抑制する場合が多い。
(注10)ブレークスルー感染
ワクチン接種後に、ウイルスに感染すること。
(注11)シュードウイルス
新型コロナウイルスのスパイクタンパク質を持たせたウイルス様粒子。細胞に一度感染できるが、増殖することはできない。
(注12)合胞体形成活性
合胞体とは、新型コロナウイルスに感染した細胞が、スパイクタンパク質を細胞表面に発現し、周囲の細胞と融合することによって形成される大きな細胞塊のこと。合胞体形成活性とは、新型コロナウイルスのスパイクタンパク質を介して、合胞体を形成する能力のこと。
問合せ先
東京大学医科学研究所 感染・免疫部門 システムウイルス学分野
教授 佐藤 佳(さとう けい)
https://www.ims.u-tokyo.ac.jp/imsut/jp/lab/ggclink/section04.html
〈報道に関する問合せ〉
東京大学医科学研究所 国際学術連携室(広報)
https://www.ims.u-tokyo.ac.jp/
北海道大学 社会共創部広報課
https://www.hokudai.ac.jp/
京都大学 渉外部広報課国際広報室
https://www.kyoto-u.ac.jp/ja
熊本大学 総務部総務課広報戦略室
https://www.kumamoto-u.ac.jp/
宮崎大学 企画総務部総務広報課
https://www.miyazaki-u.ac.jp/
HiLung株式会社 広報担当
https://www.hilung.com/