
| 発表のポイント |
|---|
|
発表概要
東京大学医科学研究所粘膜免疫学部門の神岡真理子特任研究員と清野宏特任教授らの研究チームは、千葉大学の後藤義幸准教授、倉島洋介准教授、北海道大学の中村公則准教授、綾部時芳教授などとの共同研究から糖鎖を指標としてパネート細胞を機能的に分類し、その分化や機能制御について解析したところ、腸内細菌と免疫細胞由来サイトカインがパネート細胞サブセットの分化・機能を制御していることを見出しました。パネート細胞は腸管上皮細胞の一種であり、腸管の上皮細胞層は病原微生物や有害物質に対する第一線の生体バリアとして働きます。パネート細胞は正常な腸内細菌叢の維持や病原体排除の一端を担い、パネート細胞の機能の低下は炎症性腸疾患の発症リスクになることも知られています。一方で、パネート細胞の分化および機能制御機構の理解は、正常な腸内環境の構築・維持や腸管関連疾患の新規予防・治療法の確立に繋がると考えられていますが、いまだパネート細胞制御メカニズムについては不明な点が多くあります。
研究チームは生体制御の一翼を担う糖鎖フコース(注2)に着目し、フコース転移を担う2型フコース糖転移酵素遺伝子(Fut2)のパネート細胞における発現メカニズムを解析しました。さらに腸管免疫において重要な各種サイトカイン欠損マウスを解析し、宿主免疫システムによるパネート細胞の分化・機能制御について検討しました。
その結果、パネート細胞にはFut2発現(Fut2+パネート細胞)と非発現サブセットが存在し、Fut2+パネート細胞が宿主防御に貢献するサブセットであることを見出しました。さらに、サイトカイン(注3)の中でIL-22(注4)とIL-17a(注5)がFut2+パネート細胞の分化・機能制御に関わる事も示しました。
パネート細胞は腸管関連疾患と感染症の制御に関与する事が知られており、今回見出したパネート細胞サブセットとその誘導・機能制御機構の理解は、さらなる病態形成メカニズムの解明とそれを標的とした新規予防・治療法の開発に繋がることが期待されます。
本研究成果は2022年1月13日、米国科学アカデミー紀要「Proceedings of the National Academy of Sciences of the United States of America」(PNAS、オンライン版)に掲載されました。
本研究は、日本学術振興会科学研究費助成事業 (基盤研究、若手研究、国際共同研究加速基金)、国立研究開発法人日本医療研究開発機構(AMED)の革新的先端研究開発支援事業(AMED-PRIME)、国立研究開発法人科学技術振興機構(JST)の研究成果展開事業(COI)の支援を受け実施されました。
-
発表内容
パネート細胞は腸管恒常性の維持に大きく貢献し、その分化および機能発現制御メカニズムの理解は健康的な腸内環境の構築・維持や腸管関連疾患の新規予防・治療法の確立につながる可能性を秘めていますが、いまだ不明な点が多くあります。
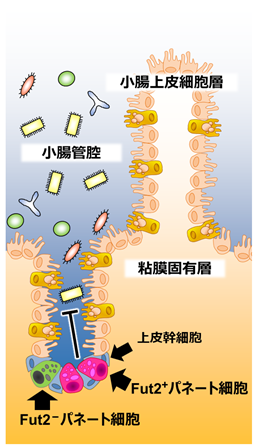
図1:小腸粘膜におけるパネート細胞の局在と、そのサブセットの発見
研究チームはこれまでに、腸内細菌との共生環境構築と感染防御に重要な腸管上皮細胞層を構成する「絨毛円柱上皮細胞」における糖鎖の付加(フコシル化)に着目し研究を展開してきました。絨毛円柱上皮細胞のフコシル化を担う2型フコース糖転移酵素遺伝子(Fut2)の発現は、自然免疫細胞の一種である3型自然リンパ球(ILC3)が腸内細菌の刺激により産生するサイトカインIL-22に誘導されることを明らかにしました。一方、腸管上皮細胞層を構成するもう一つの主要細胞である「パネート細胞」のフコシル化の生物学的意義やその発現誘導メカニズムは不明でした。そこで本研究では、パネート細胞におけるFut2発現の検討に始まりその発現メカニズムに着目し、パネート細胞の分化および機能制御メカニズムの解明を目指しました。
(緑色:パネート細胞特異的マーカーのライソザイム、赤色:フコース、黄色:細胞膜、赤矢印:フコース陽性パネート細胞、白矢印:フコース陰性パネート細胞、スケールバー:10µm)
(青色:Fut2発現、赤色:パネート細胞特異的マーカーのライソザイム、赤矢印:Fut2発現パネート細胞、白矢印:Fut2非発現パネート細胞、スケールバー:20µm)
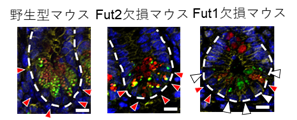
腸管においてフコシル化はFut1およびFut2が担うことから、野生型マウス、Fut1欠損マウス、およびFut2欠損マウスのパネート細胞におけるフコース発現をUEA-1レクチンを用いて免疫組織学的に観察しました(図2)。その結果、野生型マウス、およびFut2欠損マウスのすべてのパネート細胞ではフコースが観察されました。
このことから、すべてのパネート細胞のフコシル化はFut1が制御することがわかりました。しかしながら、Fut1欠損マウスにおいても一部のパネート細胞でフコースが観察されました。このことから、一部のパネート細胞ではFut1に加えてFut2もフコシル化を制御することがわかりました。
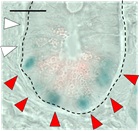
さらに、Fut2遺伝子発現を直接的に確認出来るFut2 LacZ/+マウスの小腸組織切片をLacZ染色することでパネート細胞のFut2の発現を確認し(図3)、Fut2発現パネート細胞(Fut2+パネート細胞)と非発現パネート細胞のサブセットが存在することを明らかにしました(図1-3)。
次にパネート細胞内顆粒に結合する蛍光プローブZinpyr-1とフコース特異的に結合するレクチンUEA-1を用いたフローサイトメトリー法を駆使して、パネート細胞サブセットを比較検討した結果、Fut2+パネート細胞は感染防御に重要な抗菌ペプチドを内包する細胞内顆粒を豊富に有する事から、感染防御に深く関わることが示唆されました。Fut2のパネート細胞における役割を検討する為にFut2欠損マウスと野生型マウスを比較解析したところ、Fut2欠損マウスのパネート細胞内顆粒は異常形態を示し、糞便中のパネート細胞特異的抗菌ペプチドであるαディフェンシン(注6)量が有意に減少していました。さらにマウス腸管オルガノイドを作製し、パネート細胞の顆粒分泌の様子を観察したところ、野生型マウスと比較してFut2欠損マウス由来では顆粒放出の割合が減少していました。以上の結果から、Fut2によるフコシル化はパネート細胞の顆粒分泌を制御することが示されました。
(緑色:パネート細胞特異的マーカーのライソザイム、赤色:フコース、赤矢印:Fut2+パネート細胞、白矢印:Fut2-パネート細胞、スケールバー:20µm
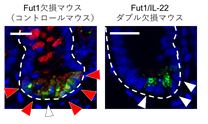
小腸におけるFut2+パネート細胞の局在を調べたところ、その多くが十二指腸ではなく回腸に局在していました。回腸には十二指腸と比較して腸内細菌が豊富に存在することを考慮し、Fut2+パネート細胞の分化と腸内細菌の関与を検討しました。腸内細菌を消失させるため抗生物質を経口投与したところ、Fut2+パネート細胞は消失しました。続いてILC3から腸内細菌依存的に産生されるサイトカインIL-22の関与を検討する為、IL-22欠損マウスを解析したところ、Fut2+パネート細胞の消失が認められました(図4)。
これらの結果からFut2+パネート細胞は腸内細菌刺激およびIL-22依存的なサブセットであることもわかりました。
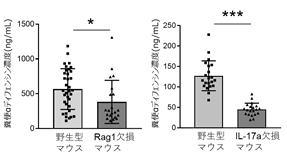
さらに宿主免疫細胞によるパネート細胞制御について詳細に検討するため、獲得免疫に関わるT細胞およびB細胞を欠損するRag1欠損マウスを解析しました。その結果、Rag1欠損マウスの糞便αディフェンシン濃度は、野生型マウスと比較して有意に減少していました。さらにIL-17a欠損マウスの糞便でも、αディフェンシンの濃度が減少していました(図5)。以上から、パネート細胞は獲得免疫細胞およびIL-17aの制御を受けることがわかりました。
発表雑誌
雑誌名:Proceedings of the National Academy of Sciences of the United States of America(PNAS、1月18日付けオンライン版)論文タイトル: Intestinal commensal microbiota and cytokines regulate Fut2+ Paneth cells for gut defense
著者:
Mariko Kamioka, Yoshiyuki Goto, Kiminori Nakamura, Yuki Yokoi, Rina Sugimoto, Shuya Ohira, Yosuke Kurashima, Shingo Umemoto, Shintaro Sato, Jun Kunisawa, Yu Takahashi, Steven E. Domino, Jean-Christophe Renauld, Susumu Nakae,Yoichiro Iwakura, Peter B. Ernst, Tokiyoshi Ayabe, Hiroshi Kiyono*(*責任著者)
DOI番号:10.1073/pnas.2115230119
アブストラクトURL:https://www.pnas.org/content/119/3/e2115230119
問い合わせ先
<研究に関するお問い合わせ>東京大学医科学研究所 粘膜免疫学部門 特任教授
清野 宏(きよの ひろし)
https://www.ims.u-tokyo.ac.jp/imsut/jp/lab/dstngprof/section02.html
<報道に関するお問い合わせ>
東京大学医科学研究所 国際学術連携室(広報)
https://www.ims.u-tokyo.ac.jp/imsut/jp/
千葉大学広報室
https://www.chiba-u.ac.jp/
北海道大学総務企画部広報課
用語解説
(注1)パネート細胞パネト細胞(Paneth cell)とも言われ、小腸上皮細胞の一種。上皮細胞層の陰窩に位置する。その主な役割は自然免疫による宿主防御であり、細胞質内に有する抗菌ペプチド αディフェンシン等を含む多数の分泌顆粒を刺激に応じて腸管管腔内に放出する。またパネート細胞は隣接する上皮幹細胞を維持するためのニッチを形成する役割も持つ。
(注2)フコース
海藻の粘り気の成分として見つかった単糖の一種。腸管においては上皮細胞に存在し、腸内細菌の栄養源として利用されることや、病原性細菌に対して防御的に働くことが知られている。
(注3)サイトカイン
免疫系細胞から分泌されるタンパク質の総称で、細胞間の情報伝達を担う。
(注4)IL-22
サイトカインの一種で、腸管においては腸管上皮細胞に作用し、腸管上皮細胞での粘液や抗菌タンパク質の産生誘導など自然免疫に関与する。
(注5)IL-17a
サイトカインの一種で、IL-22と同様に自然免疫を誘導することが知られているが、その一方で、各組織において炎症を誘導し病態を悪化させる作用も報告されている。
(注6)αディフェンシン(α-defensin, デフェンシン)
自然免疫システムで働く抗菌ペプチドの一種。ヒトやマウスをはじめとする哺乳類の消化管においては、パネート細胞の細胞内顆粒だけに発現している。細菌刺激などに応じてパネート細胞の顆粒放出により小腸内腔に分泌される。病原菌の宿主への侵入を阻止するとともに腸内細菌の組成を制御することで腸管恒常性の維持に貢献し、その異常は各種感染症をはじめ炎症性腸疾患やうつ病など様々な疾患に関係することが報告されている。