







はじめに
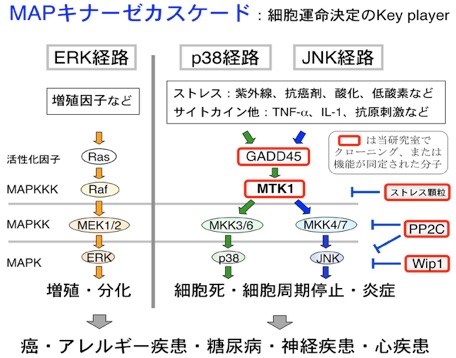 私達の研究室では、分子生物学、生化学や細胞生物学の研究手法を駆使し、細胞内情報伝達機構、特に細胞運命(増殖、分化、生存、死)の決定に最も重要なシグナル伝達システムである、MAPキナーゼ・カスケードの研究を行っています。 MAPキナーゼ・カスケードは、MAPKKK-MAPKK-MAPKという3種類の蛋白質リン酸化酵素(キナーゼ)によって構成されるシグナル伝達モジュールであり、出芽酵母からヒトに至る全ての真核生物に相同な分子が存在する細胞内情報伝達の根幹をなすシステムです。
私達の研究室では、分子生物学、生化学や細胞生物学の研究手法を駆使し、細胞内情報伝達機構、特に細胞運命(増殖、分化、生存、死)の決定に最も重要なシグナル伝達システムである、MAPキナーゼ・カスケードの研究を行っています。 MAPキナーゼ・カスケードは、MAPKKK-MAPKK-MAPKという3種類の蛋白質リン酸化酵素(キナーゼ)によって構成されるシグナル伝達モジュールであり、出芽酵母からヒトに至る全ての真核生物に相同な分子が存在する細胞内情報伝達の根幹をなすシステムです。
ヒトを代表とする哺乳類細胞には、ERK経路、p38経路、JNK経路という少なくとも3種類のMAPキナーゼ・カスケードが存在しますが、各経路の生理機能はそれぞれ異なることが知られています。
| ERK経路 | 主に増殖因子によって活性化され、細胞増殖や分化を制御する。ERK経路の上流に位置するRasや増殖因子受容体は癌遺伝子であり、この経路の異常な活性亢進が発癌を招く。 |
|---|---|
| p38およびJNK経路 | ストレス応答MAPK経路とも呼ばれる。紫外線や放射線、酸化、熱ショック、高浸透圧などの様々な環境ストレス刺激によって活性化され、ストレスを被った細胞に細胞死(アポトーシス)を誘導する。また、炎症性サイトカインや病原体の感染などによっても活性化され、免疫応答や炎症の制御に中心的な役割を果たしている。 |
これら複数のMAPキナーゼ経路が正しく制御されることで、人体の恒常性が維持されていますが、MAPK経路に何らかの制御異常が起こってしまうと、癌、アレルギー・自己免疫疾患(関節リウマチ、喘息等)、糖尿病や神経変性疾患(アルツハイマー病、パーキンソン病等)などの疾病になってしまうことが知られています。
私達は、これら難病の克服を目指してMAPK経路の活性制御機構と生理機能、および疾病との関連を分子レベルで解き明かし、さらにMAPK経路をターゲットとした分子標的薬剤を開発して疾患治療に役立てることを最終的な目標として、日々研究を推進しています。
これまでに、MAPキナーゼ経路の活性制御に関わる複数の新規遺伝子を同定することに成功し、その生理機能と疾患との関わりを明らかにして来ました。
1. 新規ヒト・ストレス応答MAPKKK、MTK1の発見と生理機能の解明
 ヒト・ストレス応答MAPキナーゼ経路の活性制御機構を解明するため、まずモデル生物である出芽酵母を用いて研究を開始し、酵母のストレス応答MAPK経路を制御する新しい2つのMAPKKK分子(Ssk2/Ssk22)、および浸透圧ストレス・センサーを同定した(Science, 1995)。
ヒト・ストレス応答MAPキナーゼ経路の活性制御機構を解明するため、まずモデル生物である出芽酵母を用いて研究を開始し、酵母のストレス応答MAPK経路を制御する新しい2つのMAPKKK分子(Ssk2/Ssk22)、および浸透圧ストレス・センサーを同定した(Science, 1995)。
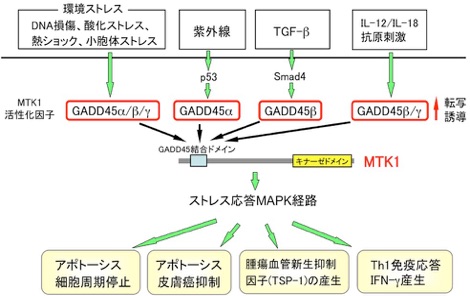
次に、この成果をヒト経路の研究へと発展させ、酵母Ssk2/Ssk22に相同な新規ヒトMAPKKK遺伝子、MTK1のクローニングに成功した。 また実際に、哺乳類細胞を用いてMTK1が、ヒト・ストレス応答MAPキナーゼ(p38およびJNK)経路を特異的に活性化するMAPKKKであることを明らかにした(EMBO J, 1997)。
様々な環境ストレス刺激が、どのようにしてMTK1を活性化するのかを明らかにするため、MTK1の制御ドメインと特異的に結合する分子のスクリーニングを行い、3種類のGADD45関連遺伝子(GADD45α/β/γ)を単離することに成功した。 さらに、これらGADD45関連分子が、様々なストレスやサイトカイン刺激によって転写誘導されるストレス応答遺伝子であり、MTK1と結合してその活性化因子として機能することを見出した。 即ち、ストレスやサイトカイン刺激によって発現誘導されたGADD45関連分子が、MTK1を介してp38およびJNK経路を活性化し、ストレスを被った細胞に死(アポトーシス)を誘導するという、新たなシグナル伝達システムの存在を示した (Cell, 1998; MCB, 2002; MCB, 2007)。
次に、GADD45-MTK1経路の生理機能の解明を推進し、まずこの経路がTGFβのシグナル伝達に関与することを見出した。TGFβは発癌抑制作用を持つサイトカインであり、実際にその下流で作用する転写因子Smad4は膵臓癌や大腸癌で高率に遺伝子変異が認められる癌抑制遺伝子である。私達は、TGFβ刺激によってSmad4依存的に発現誘導されたGADD45βが、MTK1-p38経路を活性化して腫瘍血管新生抑制因子TSP-1の発現を亢進させ、発癌阻止に作用する事を明らかにした。また、実際にSmad4に変異を持つ癌細胞では、TGFβで刺激してもGADD45βが発現しないため、MTK1-p38/JNK経路の活性化やTSP-1の発現は共に消失しており、MTK1の制御異常が発癌に関与することを示した(EMBO J, 2002, Nature Commun, 2013)。
またMTK1遺伝子欠損マウスを作成して、免疫系における機能に関しても解析を進め、GADD45分子によるMTK1-p38経路の活性化が、Th1細胞(ヘルパーT細胞の亜群)からのIFN-γ産生に必須であり、Th1免疫応答(細胞性免疫・遅延型アレルギー)の制御に中心的な役割を果たしていることを明らかにした(EMBO J, 2004)。
さらに最近、酸化ストレス環境下で、MTK1の制御ドメイン内に存在するCys残基が速やかに酸化され、その後、徐々に還元されることで酵素活性が著しく亢進し、その結果、細胞内でp38/JNK経路が強力かつ持続的に活性化して、細胞死や炎症性サイトカインの産生が誘導されることを見出した。即ち、MTK1が、生体内の酸化ストレスを感知するストレス・センサー分子として機能していることを明らかにした(Science Adv, 2020)
2. ストレス応答MAPキナーゼ経路の活性抑制メカニズムと発癌
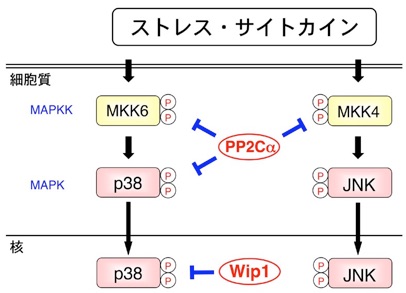 一方、ストレス応答経路の活性阻害機構に関しても研究を展開し、特にPP2C型セリン/スレオニン脱リン酸化酵素の関与を明らかにしてきた。まず、ストレス応答経路の活性化を阻害する機能を持つヒト遺伝子のスクリーニングを行い、PP2Cαがp38MAPK及びMAPKK (MKK4/6)を脱リン酸化して不活性化し、細胞のストレス応答を負に制御する分子であることを明らかにした(EMBO J, 1998)。
一方、ストレス応答経路の活性阻害機構に関しても研究を展開し、特にPP2C型セリン/スレオニン脱リン酸化酵素の関与を明らかにしてきた。まず、ストレス応答経路の活性化を阻害する機能を持つヒト遺伝子のスクリーニングを行い、PP2Cαがp38MAPK及びMAPKK (MKK4/6)を脱リン酸化して不活性化し、細胞のストレス応答を負に制御する分子であることを明らかにした(EMBO J, 1998)。
さらに、紫外線などのDNA損傷によって、p53依存的に発現誘導されるPP2C類似ホスファターゼWip1(PPM1D)が、p38やp53を脱リン酸化して、これらの分子の活性を阻害し、DNA損傷後のアポトーシスを抑制する機能を持つことを解明した(EMBO J, 2000)。
我々のこの発表を基に、Wip1はその後、様々な癌で異常な遺伝子増幅が認められる癌遺伝子であることが明らかとなった。
3. MAPキナーゼ経路のシグナル特異性決定・維持機構と疾患治療への応用
ヒト細胞にはERK/p38/JNKという、少なくとも3種類のMAPK経路が存在するが、これら複数の経路間でシグナルの誤った混線は起こらない。MAPK経路の正確なシグナル伝達を保証する機構は、細胞機能の制御に極めて重要であると考えられる。この様なメカニズムの一つとして、MAPK経路を構成する各キナーゼ分子が相互に直接結合しあう、ドッキング相互作用が重要であることが示唆されている。そこで私達は、MAPKKK-MAPKK分子間の選択的結合を規定し、シグナル伝達特異性を決定づける未知メカニズムの解明を行った。
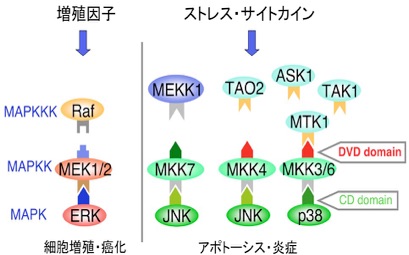 その結果、全てのMAPKK分子のC末端領域に、対応する上流のMAPKKK分子との特異的結合に必要な、新規ドッキング・サイトが存在することを見出し、DVDサイトと命名した。さらにこのDVDサイトを介した分子間相互作用が、シグナル特異性の決定、維持のみならず、MAPKKKからMAPKKへの効率的なシグナル伝達にも必須であることを明らかにした。
その結果、全てのMAPKK分子のC末端領域に、対応する上流のMAPKKK分子との特異的結合に必要な、新規ドッキング・サイトが存在することを見出し、DVDサイトと命名した。さらにこのDVDサイトを介した分子間相互作用が、シグナル特異性の決定、維持のみならず、MAPKKKからMAPKKへの効率的なシグナル伝達にも必須であることを明らかにした。
またDVDサイトと相同なアミノ酸配列を持つ人工合成ペプチドを細胞に導入して、MAPKKK-MAPKK分子間の相互作用を競合的に阻害すると、MAPKKの活性化が強く抑制される事を見出した。 即ち、ドッキング・サイトをターゲットとした分子標的薬剤を開発する事で、MAPK経路に対する新たな活性阻害剤の創薬が可能であり、癌や自己免疫疾患の治療へ応用し得ることを示した(Molecular Cell, 2005)。
4. 液-液相分離顆粒(ストレス顆粒)による、ストレス応答経路の活性阻害と抗癌剤抵抗性
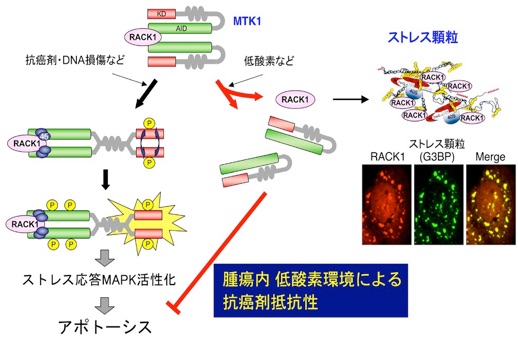 「ストレス顆粒」は、低酸素などの特定のストレス刺激によって一過性に形成される細胞質内構造体であり、その本体はmRNA、RNA結合蛋白質、及び40Sリボゾーム等からなる液-液相分離顆粒であることが知られている。しかしながらストレス顆粒形成が細胞のストレス応答に果たす役割は、ほとんど明らかにされていない。
「ストレス顆粒」は、低酸素などの特定のストレス刺激によって一過性に形成される細胞質内構造体であり、その本体はmRNA、RNA結合蛋白質、及び40Sリボゾーム等からなる液-液相分離顆粒であることが知られている。しかしながらストレス顆粒形成が細胞のストレス応答に果たす役割は、ほとんど明らかにされていない。
私達は、ストレス顆粒が、MTK1の活性化促進分子(RACK1)を顆粒内に取り込んでその機能を阻害し、MTK1−p38/JNK経路を失活させて、DNA損傷による細胞死(アポトーシス)を強く抑制する事を見出した。
ストレス顆粒の機能としては、これまでに蛋白質の翻訳抑制に作用する事のみが報告されてきたが、我々の研究によって初めて、ストレス顆粒が多彩なシグナル伝達分子とも相互作用して、細胞死誘導シグナルを負に制御する多機能構造体である事が明らかになった。
さらに我々は、この様なストレス顆粒形成による細胞死抑制が、固形癌を治療する上で問題となっている「腫瘍内部低酸素環境による癌細胞の抗癌剤抵抗性獲得」の一因となっていることを示した(Nature Cell Biology, 2008)(紹介記事)。
またストレス顆粒と神経疾患との関連についても研究を展開し、小胞体ストレスによって誘導されるストレス顆粒の形成が、酸化ストレス環境下では異常に阻害されてしまうこと、またこのことが神経変性疾患(パーキンソン病、アルツハイマー病、ポリグルタミン病等)で認められる神経細胞死の一因となることを明らかにした(Nature Commun, 2016)。
さらに最近、近接依存性プロテオミク解析技術を用いて、ストレス顆粒の構成分子を網羅的に同定することに成功するとともに、ストレス顆粒が、アポトーシス誘導に必須の実行型カスパーゼ(Casp-3/7)を顆粒内に取り込んで失活させ、様々なストレス刺激による細胞死を回避する作用をもつことを発見した(Current Biol, 2023)。
ストレス顆粒の生理機能の解明は、現在ストレス応答研究のホット・トピックとして注目を集めている。
5. ERK経路を介した発癌メカニズムの解明と診断・治療薬開発
 ERKは、様々な基質分子のリン酸化を介して遺伝子発現を調節することで、細胞増殖の制御と発癌に極めて重要な役割を果たしている。私達は、癌におけるERK経路の制御異常に関して研究を進め、これまでにMEKのSUMO化が、ERKの過剰な活性化を防いで発癌阻止に作用する事、また癌遺伝子Rasが、MEKのSUMO化を阻害してERK経路を強く活性化し、発癌を導く事を明らかにした (Nature Cell Biology, 2011)。
ERKは、様々な基質分子のリン酸化を介して遺伝子発現を調節することで、細胞増殖の制御と発癌に極めて重要な役割を果たしている。私達は、癌におけるERK経路の制御異常に関して研究を進め、これまでにMEKのSUMO化が、ERKの過剰な活性化を防いで発癌阻止に作用する事、また癌遺伝子Rasが、MEKのSUMO化を阻害してERK経路を強く活性化し、発癌を導く事を明らかにした (Nature Cell Biology, 2011)。
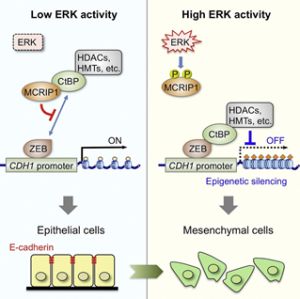
また最近、ERKによってリン酸化される基質分子を、ヒトcDNAライブラリーから網羅的に探索する新たな遺伝子スクリーニング法(酵母3-hybrid法)を開発して、これまで全く報告のない新規ERK基質分子(MCRIP1と命名)を同定することに成功した。さらに、MCRIP1の生理機能の解析を行い、MCRIP1が癌抑制遺伝子E-カドヘリンの発現制御を介して、癌の転移に重要な「上皮間葉転換(EMT)」に重要な役割を果たしていることを見出した。即ち、癌細胞では、ERKの異常な活性化によってMCRIP1が常にリン酸化された状態にあり、その結果、E-カドヘンリンの発現が低下して、癌の浸潤能・転移能が亢進していることを明らかにした(Molecular Cell, 2015)。
さらにMCRIP1遺伝子破壊マウスを作成して発生過程における機能解析も行い、MCRIP1が肺サーファクタント遺伝子(肺胞を膨らんだ状態に保つのに必要な遺伝子)の発現に必要不可欠であり、ノックアウトマウスは生後数時間以内に呼吸不全で死亡することを見出した(Commun Biol, 2019)。
 また、癌に対する新たな診断法や治療法の開発を目指して、癌細胞内で起きているシグナル伝達異常を明らかにすべく研究を推進している。これまでに、次世代シークエンサーを用いたトランスクリプトーム解析や、プロテイン・アレイ/プロテオミクス解析を実施し、癌特異的な遺伝子発現異常や翻訳後修飾異常を網羅的に同定することに成功している。
また、癌に対する新たな診断法や治療法の開発を目指して、癌細胞内で起きているシグナル伝達異常を明らかにすべく研究を推進している。これまでに、次世代シークエンサーを用いたトランスクリプトーム解析や、プロテイン・アレイ/プロテオミクス解析を実施し、癌特異的な遺伝子発現異常や翻訳後修飾異常を網羅的に同定することに成功している。
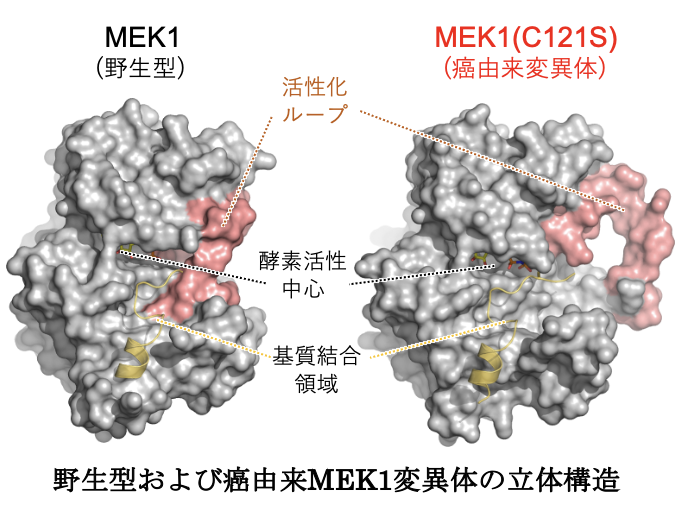
特に最近、発生異常(心肥大、精神遅延など)を伴う遺伝病(先天性Ras/MAPK症候群:RASopathy)や、癌の原因となるMEK変異体の異常活性化機構や生物学的特性を解明して、これらの分子が発生異常や発癌を導くメカニズムを明らかにした。加えてこれらの解析から、癌細胞が分子標的抗癌剤(ERK経路阻害剤)に対して抵抗性を獲得する新たな分子機構を発見するとともに、この機構を遮断して分子標的抗癌剤の効果を高める新たな癌治療法を提唱した(Nature Commun. 2022a)。
さらに、ERK経路が細胞増殖や発癌を導く新たなメカニズムとして、増殖刺激や癌遺伝子によって活性化したERKが、RNAポリメラーゼII(Pol-II)の転写伸長反応を促進させる作用をもつこと、またその結果、初期応答遺伝子群(Fos、Junなど、細胞増殖に重要な遺伝子)のmRNA合成が亢進して、細胞増殖や癌の更なる悪性化が惹起されていることを発見した(Nature Commun. 2022b)
現在、これらの基礎データを活用して癌に対する新たな分子診断法や分子標的治療薬の開発を進めている。
6. シグナル伝達分子の時空間動態変化による生命機能制御機構の解明
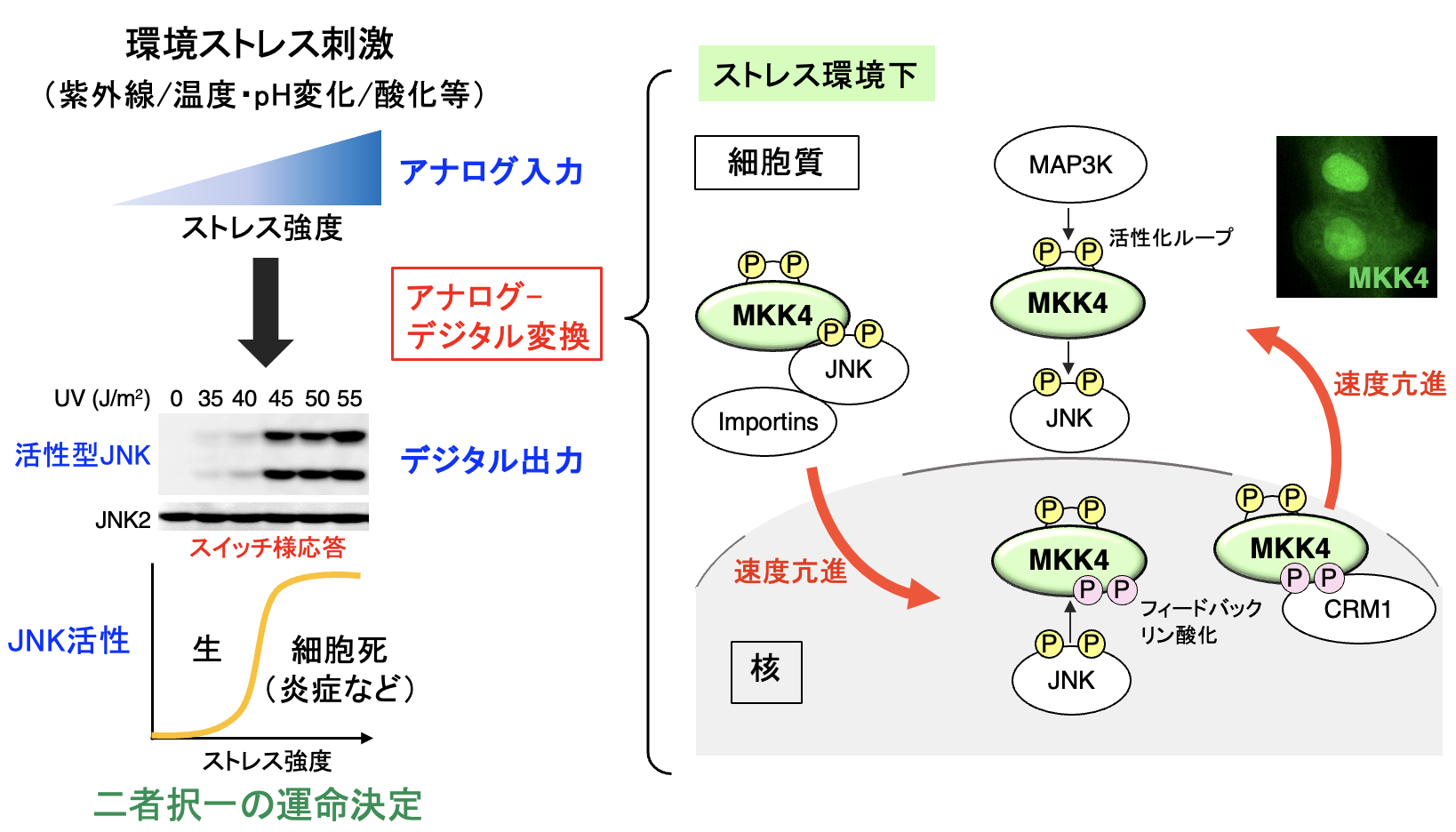
人体は、常に様々な環境ストレス(紫外線、温度/pH変化など)に曝されている。この様な刺激に対して、細胞が生存し得る至適条件は限られた範囲内であり、一定以上の(閾値を超えた)強度のストレスに曝された細胞は細胞死により排除される。即ち、個々の細胞においては、その強度が連続的に変化する「S/N比の低いストレス刺激」(アナログ入力)から、「生きるか死ぬか」という二者択一の運命決定(デジタル出力)が導き出されている。この様なストレス刺激に対する細胞内情報伝達の「アナログ-デジタル変換」は、生物が環境変化に適応する上で必須であるにも関わらず、そのメカニズムは殆ど明らかにされていない。
私達はこの未知の機構を解明すべく、ストレス下での細胞運命決定に重要なJNK経路について詳細な解析を実施した。その結果、JNKの上流因子であるMKK4が、無刺激状態で核と細胞質の間を緩やかに往復(シャトル)していることを見出すとともに、ストレス環境下では、その速度が著しく(100倍以上)亢進することを発見した。さらに、このMKK4のストレス依存的な動態変化そのものが、スイッチ様のJNK活性化パターンを生み出す原動力であり、アナログ入力であるストレス刺激からデジタル出力であるJNK応答を導く「アナログ-デジタル変換器」として機能していることを解明した。実際に、MKK4の動態変化を人工的に破綻させると、JNKがスイッチ様応答から、刺激強度に比例した線形応答に変化してしまい、低線量の紫外線に対しても活性化する様になって、細胞死や炎症が誘発されることを見出した。即ち、シグナル伝達分子の「動き」そのものが情報となって、環境変化に対する生体応答を精緻に制御するという、シグナル伝達の新たな制御原理を明らかにした(Nature Commun, 2026)。
おわりに
これまで私達は、主にMAPK(ERK/p38/JNK)経路の活性制御機構の研究を推進し、この経路が細胞運命決定や、免疫応答の制御に極めて重要であることを示すと共に、その制御異常が発癌に関与することを明らかにしてきました。さらに最近、臨床検体を用いたゲノム・ワイドの解析により、様々な癌で見出されるMKK4(ストレス応答経路のMAPKK)の機能欠損変異が、発癌プロセスに極めて重要な遺伝子変異(driver mutation)の一つであることや、p38経路の活性化がoncogene addiction(癌遺伝子の不活性化によって誘導される癌細胞特異的なアポトーシス)に最も重要であり、癌遺伝子をターゲットとした分子標的治療において、癌細胞の排除に中心的な役割を果たすシグナル伝達システムであることなどが相次いで報告されています。
また現在、多くの製薬企業によってMAPK経路の選択的阻害剤が開発され、臨床試験が開始されており、癌、関節リウマチ、神経変性疾患や2型糖尿病などに対する新たな分子標的治療薬としての可能性が大きく注目されています。 しかしながら、MAPキナーゼ・カスケードの活性制御機構や生理機能には、未だ不明な点が数多く残されており、また疾患における制御異常の詳細も明らかにされていません。これらの問題の解明は生物学的に極めて重要であるばかりでなく、癌、アレルギー・自己免疫疾患、神経変性疾患や糖尿病などを始めとする様々な難治性疾患の病因・病態の理解と、新たな分子標的治療薬開発の観点からも必要不可欠です。
私達は、細胞内シグナル伝達機構の基礎研究を通して、疾患の病因・病態を分子レベルで解き明かすと共に、新規治療法の開発に繋がる応用研究を積極的に展開して行きたいと考えています。