

研究内容
1.はじめに
さまざまな感染症を引き起こすウイルスの正体は、DNAまたはRNAのいずれかを遺伝子として保有し、そのまわりをタンパク質の外殻が囲んでいる極めて小さく、かつ、単純な構造をもった粒子です。興味深いことに、この小さく単純なウイルスがヒトや動物に感染すると、複雑な病気を引き起こします。そして、これら感染症の制圧は困難なものが多いのが現状です。
2.単純ヘルペスウイルス
当研究室では、DNAを遺伝子として持つウイルスのプロトタイプである「単純ヘルペスウイルス」をモデルとし、最先端かつ多面的なウイルス研究を推進しています。単純ヘルペスウイルスは、ヒトに脳炎、粘膜疾患、皮膚疾患といった多様な病態を引き起こし、全世界で数多の人々が罹患する医学上極めて重要なウイルスです。抗ウイルス薬が開発されているものの、一部の疾患にはその効果は限定的であり、ワクチンは開発されていません。よって、全世界的なアンメットメディカルニーズ(未充足な医療ニーズ)が高いウイルス感染症です。また、単純ヘルペスウイルス感染症は、エイズの原因ウイルスであるヒト免疫不全ウイルスの感染危険度を2~4倍程度増加させ、認知症の増悪に関与することも報告されています。
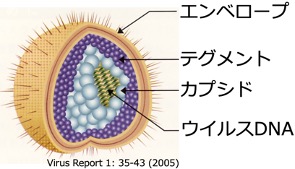
単純ヘルペスウイルスは、様々な培養細胞で効率的に増殖し、多様なヒトの病態を容易に再現できる小動物モデルが存在します。また、古くから(約100年前)から精力的に研究が推進され、多くの研究知見の蓄積があることから、ウイルス学で最も研究が進展しているウイルスの1つです。実は、ヒトの病態を容易に再現できる小動物モデルが存在し、研究が進展しているヒト病原性ウイルスは意外にも少ないのです。単純ヘルペスウイルスをモデルとすることにより、最先端かつ多面的なウイルス研究を推進することが可能となります。
3.研究の進め方
研究を推進するには、実験を遂行するための技術基盤が必須です。最先端研究の現場では、既存の技術基盤では不十分で、自ら新しい技術基盤を開発しなければならない場合が少なくありません。当研究室では、このような新しい技術基盤の開発を重要視しており、これまでに、ウイルス改変系、ウイルス遺伝子解読系、ウイルス特異酵素の解析系、生きた感染細胞でのリアルタイムイメージング系といった新しい技術基盤を開発してきました。
これら独自の技術基盤を駆使し、ウイルス学分野以外の様々な研究分野の研究者と緊密かつ実効性のある共同研究を行っています。そして、試験管内での生化学的解析および分子生物学的解析から、培養細胞レベルでの細胞生物学的解析、ウイルス学的解析、免疫学的解析、オミックス解析に加え、病態モデル動物や遺伝子改変動物を用いた生体レベルの解析やヒトサンプルを用いた解析、さらには、情報科学的解析等、様々な分野の最先端の解析手法をフルに活用し、多分野融合的なウイルス研究を推進しています。
4.最先端基礎研究とそれに基づく橋渡し研究
「ウイルスがどのように増殖するか?」「どのように宿主に病気を引き起こすか?」といったウイルス学の根幹をなす命題に迫るべく、また、研究成果を新しいウイルス制御法の開発に繋げるため、戦略的な最先端基礎研究を推進しています。ウイルスの増殖・病態発現機構の全体像は、細胞侵入、遺伝子発現、ゲノム複製、細胞内輸送、粒子形成、免疫応答、病原性といった複雑な感染現象の総和です。当研究室では、研究室のスケールメリットと多分野融合的研究を最大限活用することによって、様々な感染現象を統合的に解析し、ウイルスの増殖・病態発現機構の全体像に迫ることを目指しています。
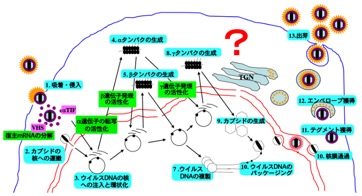
これら基礎研究で得られた独自の知見を基に、ウイルスの制圧に直結する新しいワクチンや抗ウイルス剤の開発に繋げる橋渡し研究を行っています。これまでに、当研究室の研究シーズが様々な国内外の企業に導出されています。また、導出先の国内企業の研究者が当研究室に常駐し、共同開発を行った実績もあります。
5.一般の生物研究では解明しえない細胞・生理制御機構を紐解く研究
歴史的に、スプライシング、癌遺伝子、mRNAのキャップ構造、細胞融合など、ウイルス研究から発見された重要な細胞機構は数多くありますが、現在では、ウイルス研究から新しい細胞機構が明らかにされることはあまり無くなりました。一方で、宿主細胞無しでは生存できないウイルスは、通常は眠っている細胞機構や一般の生物研究では見つけにくい細胞機構を活性化させて利用(ハイジャック)することが明らかになってきました。当研究室では、ウイルスと宿主細胞の相互作用を精緻に解析することによって、一般の生物研究では見過ごされてしまう未知な細胞機構を解き明かす研究を推進しています。既に当研究室では、ウイルス研究から、生物学上極めてユニークな細胞における核外輸送の分子機構とその意義を明らかにしただけでなく、その分子機構がウイルス感染症とは全く関連の無い遺伝病に関与していることを明らかにしています(下記、プレスリリースv参照)。
6.ウイルス自体を生体恒常性因子として捉え直しその意義を解明する研究
単純ヘルペスウイルスが属するヘルペスウイルス科のウイルスは、宿主に潜伏感染と回帰発症(再発)を長い間繰り返します。実は、宿主の一生において、ヘルペスウイルスが病気を引き起こす期間は極めて短く、大部分の期間は病気を引き起こさない潜伏感染として宿主と共存します。興味深いことに、ヘルペスウイルスの潜伏感染が、宿主の細菌感染や癌に対する抵抗性などを制御していることが近年明らかになってきました。つまり、ヘルペスウイルスは宿主に病気を起こすという負の因子であるだけでなく、宿主の生命活動に寄与する正の因子であるという新しい概念が生まれつつあります。当研究室では、ウイルスを生体恒常性因子として捉え直し、その意義を解明するという次世代ウイルス学にも挑戦しています。
7.当研究室の研究内容がわかる日本語総説
- ヘルペス脳炎を司る新規内因性免疫回避機構の解明 ウイルス 第75巻 第2号 (2025年)
- 非標準的ウイルス遺伝子解読法の確立による新規ヘルペス脳炎制御因子の同定 ウイルス 第70巻 第2号(2020年)
- 単純ヘルペスウイルス 〜基礎研究,最近の進展〜 ウイルス 第68巻 第2号(2018年)
- 単純ヘルペスウイルスの増殖・病態発現機構の解明 第54回小島三郎記念文化賞(2019年)
- 単純ヘルペスウイルス1型のコードするVP22はAIM2インフラマソームを阻害し生体における増殖に貢献する
- ライフサイエンス新着論文レビュー FIRST AUTHOR'S(2018年)
- 単純ヘルペスウイルス(HSV) ウイルス 第60巻 第2号(2010年)
- 生きた細胞におけるウイルスの可視化 ウイルス 第58巻 第2号(2008年)
- BACシステム:大腸菌遺伝学とウイルス学が融合した新しいヘルペスウイルスの遺伝子改変法 ウイルス 第54巻 第2号(2004年)