1.はじめに
2.ヘルペスウイルスとは?
3.ヘルペスウイルス新規受容体の発見
4.ウイルス粒子およびその成熟過程の可視化
5.大腸菌遺伝学とウイルス学の融合:新しいウイルス改変系の確立
6.ヘルペスウイルスがコードするプロテインキナーゼ
1.はじめに
近年、ウイルス病がしばしば世間を騒がす。これらの感染症を引き起こすウイルスの正体は、DNAまたはRNAのいずれかを遺伝子として保有し、そのまわりをタンパク質の外殻が囲んでいる極めて小さく、かつ、単純な構造をもった粒子である。興味深いことに、この小さく・単純なウイルスがヒトや動物に感染すると、複雑な病気を引き起こす。そして、これら感染症の制圧は困難なものが多いのが現状である。
本研究室では、DNAを遺伝子として持つウイルスであるヘルペスウイルスを研究対象とし、ウイルス感染症に対する制御法開発を最終目標にして、研究を推進している。
具体的には、独自に開発したウイルス遺伝子操作系、ウイルス特異酵素の解析系、生きた感染細胞でのリアルタイムイメージング系を駆使し、試験管内での生化学的および分子生物学的解析から、培養細胞レベルでの細胞生物学的およびウイルス学的解析、さらには、病態モデル動物を用いた実験動物学的解析等、多岐に渡る研究を行っている。そして、医学上極めて重要なウイルスの病原性発現機構の全体像を明らかにし、それを基に、新しいウイルス制御法の確立、さらには調教ウイルスによるヒト疾患治療への応用という命題にチャレンジしている。
2.ヘルペスウイルスとは?
ヘルペスウイルス目に属するウイルスの総称をヘルペスウイルスと呼ぶが、ヘルペスウイルスは牡蠣などの無脊椎動物から、魚類、鳥類、ほ乳類などを含む脊椎動物に至るまで幅広く分布しており、現在までに350種類以上のヘルペスウイルスが、様々な種から発見されている。
ヒトを宿主とするヘルペスウイルスは8種類あり、神経疾患、粘膜性疾患、皮膚疾患、腫瘍性疾患といった様々な病態を引き起こす。また、馬、牛、豚などの家畜や、犬、猫などの伴侶動物には、それぞれ固有のヘルペスウイルスがあり、宿主に重篤な病気を引き起こす。さらに、コイヘルペスウイルスや牡蠣ヘルペスウイルス感染は養殖業に多大な経済的損失を与える。このように、ヘルペスウイルスは、医学・獣医学・畜産・水産といった多領域において極めて重要な病原体群といえる。
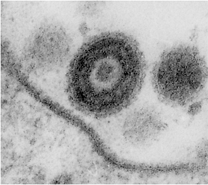
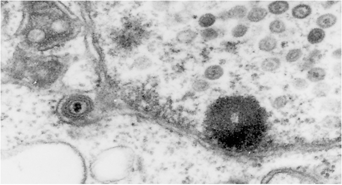
細胞外の成熟ヘルペスウイルス粒子 核内のカプシドおよび核膜を通過中のウイルス粒子
提供:板垣慎一博士
単純ヘルペスウイルス (HSV: herpes simplex virus)
HSVは、数あるヘルペスウイルス中で、最も研究が進んでいることより、ヘルペスウイルスのプロトタイプと呼ばれている。HSVは、ヒトに脳炎、性器ヘルペス、皮膚疾患、眼疾患、小児ヘルペスなど、多様な病態を引き起こす。脳炎は特に恐ろしく、無治療での致死率は70~90%と非常に高い。ノーベル賞の受賞対象である抗ヘルペスウイルス剤・アシクロビルの投与で、致死率は10%程度に低下する。しかし、生存した患者の2/3には中および重度の後遺症が残る。また、性病としてのHSVの重要性は高く、アメリカ合衆国では、年間約50万人が性器ヘルペスの初感染に罹り、約1,000万人が再発性の性器ヘルペスで苦しむ。HSV感染症の医療費は、年間30億ドル(日本円にして3,500億円)と試算されており、HSV研究の重要性は明らかである。また、HSVは強い神経指向性や高い殺腫瘍能力を有することより、遺伝子治療やウイルス療法の分野でも医学的利用が試みられている。
本研究室では、ヘルペスウイルスの中でも最も研究が進んでおり、その研究成果が多くの他のヘルペスウイルス研究に効率的にフィードバックされているHSVをモデルとし、そのウイルス増殖機構・病態発現機構を分子から個体レベルで解明することによって、ヘルペスウイルス感染症に対する普遍的な新規制御法を開発することを目指し、戦略的な基礎研究を推進している。
3.ヘルペスウイルス新規受容体の発見
ウイルスはウイルス粒子上にある糖タンパク質が細胞膜上にある特異的な受容体と会合することによって細胞内に侵入する。これまでの研究からHSVの細胞侵入にはウイルス粒子上にある糖タンパク質Bおよび糖タンパク質Dがそれぞれ異なる受容体と会合することが必要であると考えられてきた。しかし、生体内でのウイルス感染に主要な役割を果たすgD受容体は明らかになっている一方で、糖タンパク質Bに対する受容体は不明であった。
我々は、HSVの糖タンパク質Bが非筋肉ミオシンIIA (NM-IIA: non-muscle myosin IIA)の重鎖(NMHC-IIA: non-muscle heavy chain IIA)と会合することにより、ウイルスが細胞内に侵入することを発見した。興味深いことに、受容体は通常、細胞膜表面に発現していることが知られているが、NM-IIAは細胞表面にはそれほど発現しておらず、ウイルスの侵入開始に伴い速やかに細胞表面に誘導され受容体として機能することが明らかとなった。このユニークなNM-IIAの細胞表面への誘導は、細胞のタンパク質リン酸化酵素であるミオシン軽鎖キナーゼ(MLCK: myosin light chain kinase)によるNM-IIAの制御軽鎖(RLC: regulatory light chain)のリン酸化によって引き起こされており、ミオシン軽鎖キナーゼの特異的阻害剤であるML-7はNM-IIAの細胞表面への誘導を抑制し、単純ヘルペスウイルス感染を阻害できることが明らかになった。さらに、マウス角膜炎モデルにおいて、ML7を感染前にマウスに点眼すると、マウスの角膜炎症状および致死率が顕著に低減した。これらの結果は、「HSVがNM-IIAを速やかに細胞表面に誘導し、受容体として利用することによって細胞に侵入する」という全く新しい感染機構を解明したという点で学術的に高い意義を有するだけでなく、NM-IIAおよびその制御機構を標的としてHSV感染症に対する新薬を開発できる可能性を示している。本研究はNatureに掲載され、国内新聞各紙で報道された。
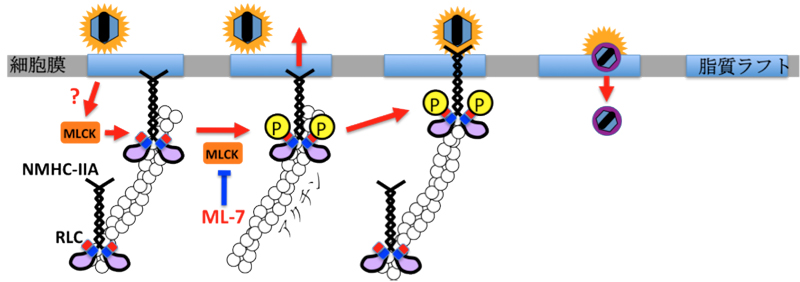
ウイルス感染症の制御法で最も効果的であるのはワクチンであることは論を待たない。しかし、大部分のヘルペスウイルス感染症のように、効果的なワクチン開発が困難であるウイルス感染症の制御は、抗ウイルス剤に頼らなければならないのが現状である。細菌に対する抗生物質を鑑みれば明らかなように、抗微生物薬の標的は、微生物に特異的な現象でなければならない。さもないと、副作用が問題となる。しかし、ヘルペスウイルスの宿主細胞依存性は極めて高く、ウイルス特異的な現象は限られている。それでは、ウイルス特異的な現象とは何であろうか?1つはウイルス特異酵素が司る感染現象である(下記5参照)。もう1つは、ウイルス粒子そのもののおよびその成熟過程であると言える。つまり、ウイルス粒子の成熟過程はウイルス特異的な現象として、新しい抗ウイルス戦略構築の有力な標的となりうる。
ウイルス粒子成熟過程の解析では、ウイルス粒子を観察することが必須である。しかし、ウイルス粒子を観察するためには、電子顕微鏡が必要であった。電子顕微鏡の解析では、感染細胞を固定する必要がある。しかし、ヘルペスウイルス粒子成熟過程は極めてダイナミックであることより、固定した細胞から得られる情報は限られていた。近年の光学顕微鏡の技術的進歩、また、様々な蛍光蛋白質および蛍光物質の開発は、生きた細胞内のウイルス粒子を光学顕微鏡で観察することを可能としつつある。
ヘルペスウイルス粒子は、3つ(カプシド、テグメント、エンベロープ)のタンパク質コンポーネントから構成されている。我々は、HSV粒子の3つ全てのコンポーネントを異なる蛍光蛋白質で標識した組み換えウイルス(YK608)の作製に成功した。重要な点は、YK608が野生体のウイルスとほぼ同様な増殖能を有し、かつ、ウイルス1粒子レベルの観察が可能なことである。つまり、作製された組み換えウイルスを用いれば、ウイルスの侵入からウイルス粒子成熟までの一連のウイルス生活環を可視化することが可能である。本研究はNHKサイエンスゼロで紹介された。
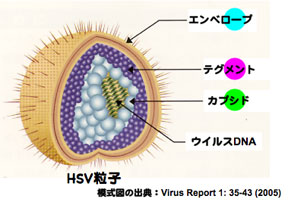
3つのコンポーネントが異なる蛍光蛋白質で標識されている組み換えウイルスYK608の模式図。
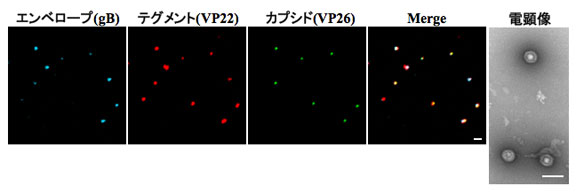
精製ウイルスを共焦点レーザー顕微鏡で観察すると、3色の蛍光を発するウイルス粒子が観察される。
さらに我々は、HSV感染によって、カプシド、テグメント、エンベロープが集積する新しいウイルス粒子成熟の‘場’が、細胞の基底面に多数形成されることを明らかにした。HSV感染によってゴルジ関連オルガネラが細胞質全体に拡散することが知られていたが、この我々が見出したウイルス粒子成熟の‘場’に、HSV感染によって細胞質に拡散したトランスゴルジネットワーク(TGN: Trans-Golgi Network)のマーカーが特異的にリクルートされていることが明らかになった。つまり、HSV粒子最終成熟の‘場’は、TGNであり、HSVは複数のウイルス成熟の‘場’を創り出すためにTGNを拡散させることが示唆された。
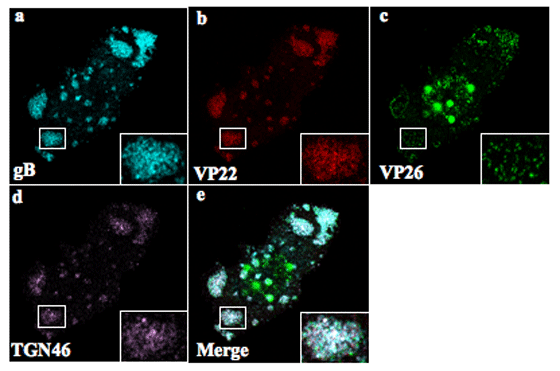
YK608感染細胞の基底面には、3つのウイルスコンポーネントが集積するウイルス成熟の’場’が多数観察される。
その’場’にTGNのマーカーが特異的にリクルートされている。
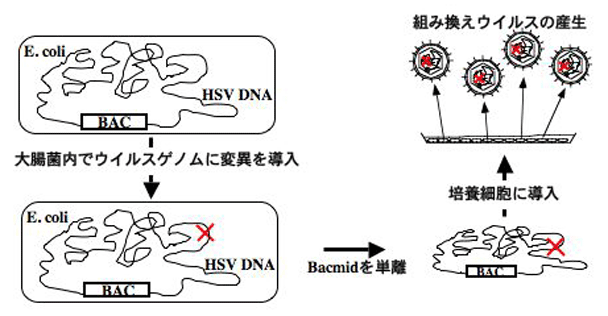
5.大腸菌遺伝学とウイルス学の融合:新しいHSV改変系の確立
HSV研究やHSVの医学的利用において、ウイルス改変系は極めて重要な技術である。ウイルスの病原性発現機構や増殖機構の解析には、標的ウイルス因子に改変を施した変異ウイルスの作製が、また、遺伝子治療ベクターの開発や改良には、病原性因子の不活化や外来遺伝子の搭載等が必須になってくる。HSV のウイルス改変法は、約四半世紀も前に確立されていた。しかし、約150kbpの巨大なウイルスゲノムを有するHSVの遺伝子操作過程は煩雑であり、作製に長期間と熟練を要した。我々は、(i) 完全長のHSVゲノムをBAC (bacterial artificial chromosome) にクローニングし大腸菌に保持させ、(ii) 大腸菌の遺伝学を利用してウイルスゲノムに変異を導入し、(iii) 変異ゲノムを培養細胞に導入することによって、(iv) 変異ウイルスを再構築させる、新しいHSV改変系を確立した。このウイルス改変系は、従来法に比して、著しく簡便であり、作製に平均2〜3ヶ月要したノックアウトウイルスの作製が、1週間から10日で作製可能となった。さらに重要なことは、大腸菌に保持された完全長のHSVゲノムから再構築された組み換えウイルスがin vitroおよびin vivoにおいて野生体の性状を呈することである。「全ての組み換えウイルス・遺伝子治療ベクターは野生体に由来する」という事実を鑑むと、我々が確立した系は、あらゆる遺伝子治療ベクターおよび組み換え変異ウイルスの作製に利用可能である。本系は、日本はもとより世界各国のHSV研究者、遺伝子治療の研究者に分与され、HSVの研究およびHSVベクター普及に貢献している。
6.ヘルペスウイルスがコードするプロテインキナーゼ
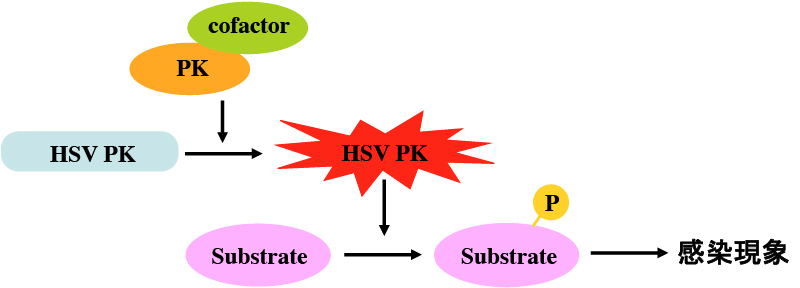
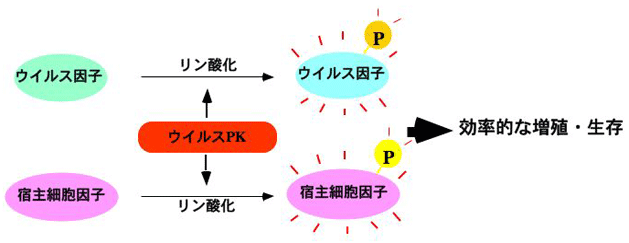
ウイルスは宿主細胞なしでは生存することができない。したがって、自身のウイルス因子を利用して、様々な宿主細胞機構を制御する(のっとる)ことによりその生存を図っている。興味深いことに、ヘルペスウイルス、ポックスウイルス、ロタウイルスはPK(プロテインキナーゼ)をコードしている。周知のように、PKによるタンパク質のリン酸化は、標的タンパク質の活性制御を司る最も一般的な修飾であり、様々な細胞機構(転写、翻訳、細胞周期、タンパク分解系、アポトーシス、etc.)がリン酸化によって制御されている。このように、様々な細胞機構を制御しうるPKをウイルスが保持していることは、宿主細胞をのっとるためには好都合である。つまり、ウイルスは自身のPKによってウイルス因子や宿主細胞因子をリン酸化し、それらの活性を制御することによって、ウイルスの増殖や生存に有利な環境を創り出していることが想像される。また、ウイルスPKはウイルス特異酵素であることより、新しい抗ウイルス剤の理想的な標的であるともいえる。このように、ウイルスPKはウイルス増殖機構や抗ウイルス戦略を考える上で魅力的な研究対象である。
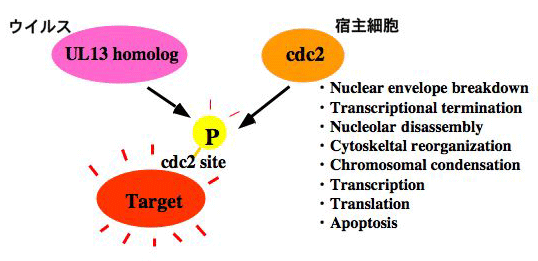
ウイルスPKの機能発現機構解明の第1ステップは、標的基質を同定することにある。しかし、ウイルスPKに対するウイルス側の標的因子が次々に同定されたのに対し、宿主細胞標的因子に関わる知見は皆無であった。我々は、HSVがコードするPKであるUL13が宿主翻訳因子EF-1 deltaをリン酸化することを明らかにした。本知見は、ウイルス特異PKの宿主細胞標的因子を、ウイルス学ではじめて同定したものとなった。また、その解析途中で偶然、UL13と宿主細胞PKである細胞周期依存PK (cdks)が同一アミノ酸をリン酸化することを見出し、UL13の機能が、宿主細胞PKであるcdksの模倣であることが明らかになった。Cdksは、転写、翻訳、細胞骨格、核膜、クロマチンといった様々な細胞機構を制御している。Cdks様の活性をもつPKをウイルスが保持することは、「宿主細胞機構をのっとる」というウイルスの目的遂行のためには、大きなメリットであると考えられる。
PKは、コファクターとの相互作用やリン酸化等によって活性化された後、基質をリン酸化することによってその機能を発揮する。HSV PKはウイルス病原性に寄与することが明らかになっているが、その活性化機構やウイルス病原性発現に関与する基質は不明であった。我々は、HSV PKの1つであるUs3の活性化が自己リン酸化によって極めて厳密に制御されていることを明らかにし、その活性制御はウイルス病原性発現に大きな役割を果たしていることを明らかにした。また、ウイルス病原性発現に寄与するUs3 PKの基質を複数同定し、ウイルス病原性発現の分子機構を明らかにした。