研究内容
TLR3、TLR7、TLR9による核酸認識機構および活性制御機構
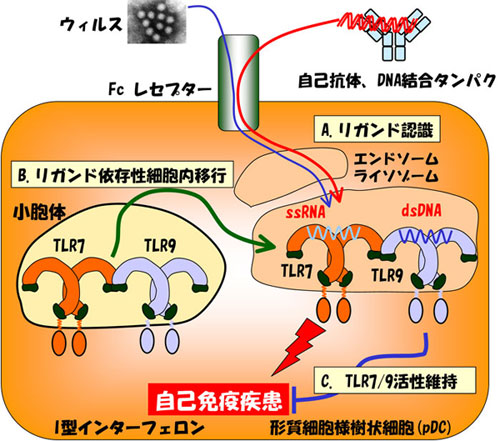
核酸を認識するTLRは、誤って自己に応答する危険性をかかえています。実際SLEなどの自己免疫疾患において、樹状細胞に発現するTLR7、TLR9によるI型インターフェロン(IFN)産生誘導が、病態形成に関与することが明らかにされつつあります。自己由来の核酸は、通常迅速に分解されるためTLR7やTLR9には認識されませんが、核酸に対する自己抗体や、LL37やHMGB1などのDNA結合タンパクと複合体を形成すると安定化し、TLR7/9を。活性化しうるようになります。TLR7/9は小胞体に局在しますが、活性化にともない、エンドソーム・ライソソームに移行し、核酸を認識します。核酸タンパク複合体はTLR7/9の細胞内局在にも影響します(図)。このように、TLR7/9による核酸認識には、核酸に結合するタンパクやTLR7/9の細胞内移行など多くのステップを必要とします。これらのメカニズムを解明し、さらに自己免疫疾患におけるその異常を検討することを目指しています。
- A.TLR7/9のリガンド認識におけるカテプシンの役割
- TLR7/9による核酸認識には、多くの分子が関与している可能性があります。そこで、そのような分子を検索するために、TLR7/9を発現していても、NF-kBの活性化が認められない細胞株を見出し、NF-kBの活性化をGFP(Green Fluorescent Protein)の発現誘導で検出できる細胞株を確立しました。この細胞に、TLR7/9に応答するマクロファージ細胞株RAW264からのcDNAを導入し、TLR9の応答性を獲得させるcDNAを単離しました。その結果、カテプシンB、カテプシンLを同定しました。カテプシンB/Lはエンドソーム・ライソゾームに分布するプロテアーゼで、TLR9のDNA認識に如何に関わっているか検討を進めています。
- B.TLR7/9のリガンド依存性細胞内移行の解析
- TLR7/9のリガンド依存性細胞内移行には、Unc93B1が重要な役割をしていることが最近報告されました。我々がクローニングした、TLRに対しシャペロン活性を持つPRAT4Aをノックダウンした場合にも、TLR9は小胞体からエンドソーム・ライソゾームへの移行が障害されます。PRAT4AやUnc93Bが如何にしてTLR7/9の細胞内移行を制御しているのか、検討しています。
- C. TLR7とTLR9の応答性の制御機構
- さらに興味深いことに、TLR7とTLR9は類似した構造を持ち、ほぼ同じシグナル伝達分子を活性化すると考えられているにもかかわらず、自己免疫疾患の病態へはむしろ相反する方向で関与している可能性が示唆されています(図)。自己免疫疾患モデルマウスMRL/lprにおいて、TLR7欠損により症状が緩和する一方、TLR7の過剰発現によって逆に自己免疫疾患となることがTLR7トランスジェニックマウスおよびミュータントマウスYaaによって示されています。これと対照的に、TLR9が欠損すると、MRL/lpr、PLCγ(Phospholipase Cγ) 遺伝子欠損マウスなどで見られる自己免疫疾患が逆に悪化することがわかっています。TLR7とTLR9には相反する機能が有り、双方を発現する樹状細胞においてはTLR7とTLR9の活性を同時に制御する必要があると推測されます。現在、我々はその機構についての解析を進めています。