2. 細胞老化維持機構
老化細胞は、恒久的な増殖停止を特徴とする細胞応答である。老化細胞は顕微鏡下で観察すると、若い細胞に比較して 扁平・肥大化しており、また細胞質に液胞が発達している。このような形態的特徴以外に、老化細胞は恒久的細胞周期停止 のために特異的にCdk阻害たんぱく質p16の発現が上昇している。さらに、老化細胞は様々な炎症性サイトカインや 増殖因子を分泌しており、これをSenescence Associated SecretoryPhenotype(SASP)と呼んでいる。 SASPがどのような生理的意義を持っているのかまだ分かっていないが、組織微小環境に何らかの影響を与えていることは容易に 推察できる。近年、SASPが誘導する組織微小環境における慢性炎症場ががん発症の母地になっている可能性が示唆され 始めている。以上の事を考え合わせると、老化細胞におけるSASP誘導や、p16の発現制御機構を解明することは、 老化細胞の維持のみならず、がんを含めた様々な加齢性疾患の病態を考える上で重要と思われる。
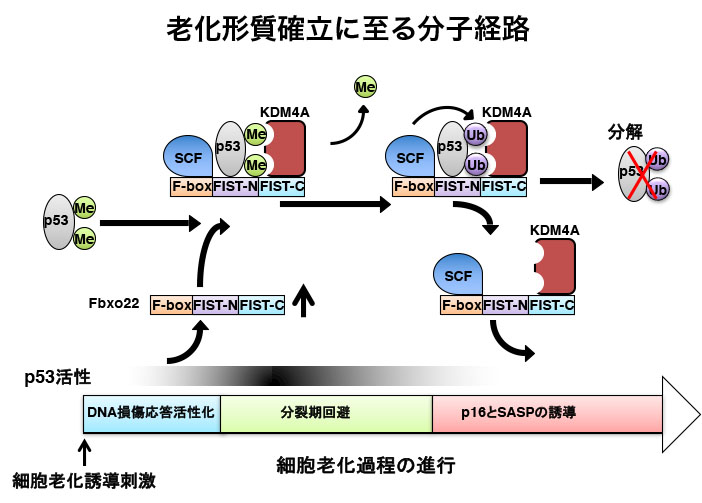
これまで、SASPの誘導やp16の発現上昇に、p53が抑制的に機能していることが知られていた。このことは、細胞老化維持過程においては、 誘導過程で活性化されたp53が不活性化される必要があることを示している。我々は、老化細胞特異的に発現する因子が、 老化細胞維持においてp53を負に制御していると考え、この因子の同定を試みた。 具体的には、老化細胞が扁平で肥大化していることを利用して細胞を選別し、老化細胞特異的に発現している遺伝子群を同定した。 その結果、Fbxo22というF-boxたんぱく質遺伝子が老化細胞特異的に発現誘導されていることが分かった。 Fbxo22は SCFたんぱく質分解複合体と結合し、p53をユビキチン化・分解誘導することが示され、老化細胞におけるSASPやp16発現 と関連していると考えられた。興味深いことに、Fbxo22はp53と同時にKDM4Aリジン脱メチル化酵素とも結合し、 SCFと合わせて大きな複合体を形成していることが分かった。このことから、SCF-Fbxo22-KDM4A複合体はメチル化p53を 標的にしている可能性が示唆された。実際、Fbxo22の発現を抑制すると、メチル化p53が特異的に増加することも示された。
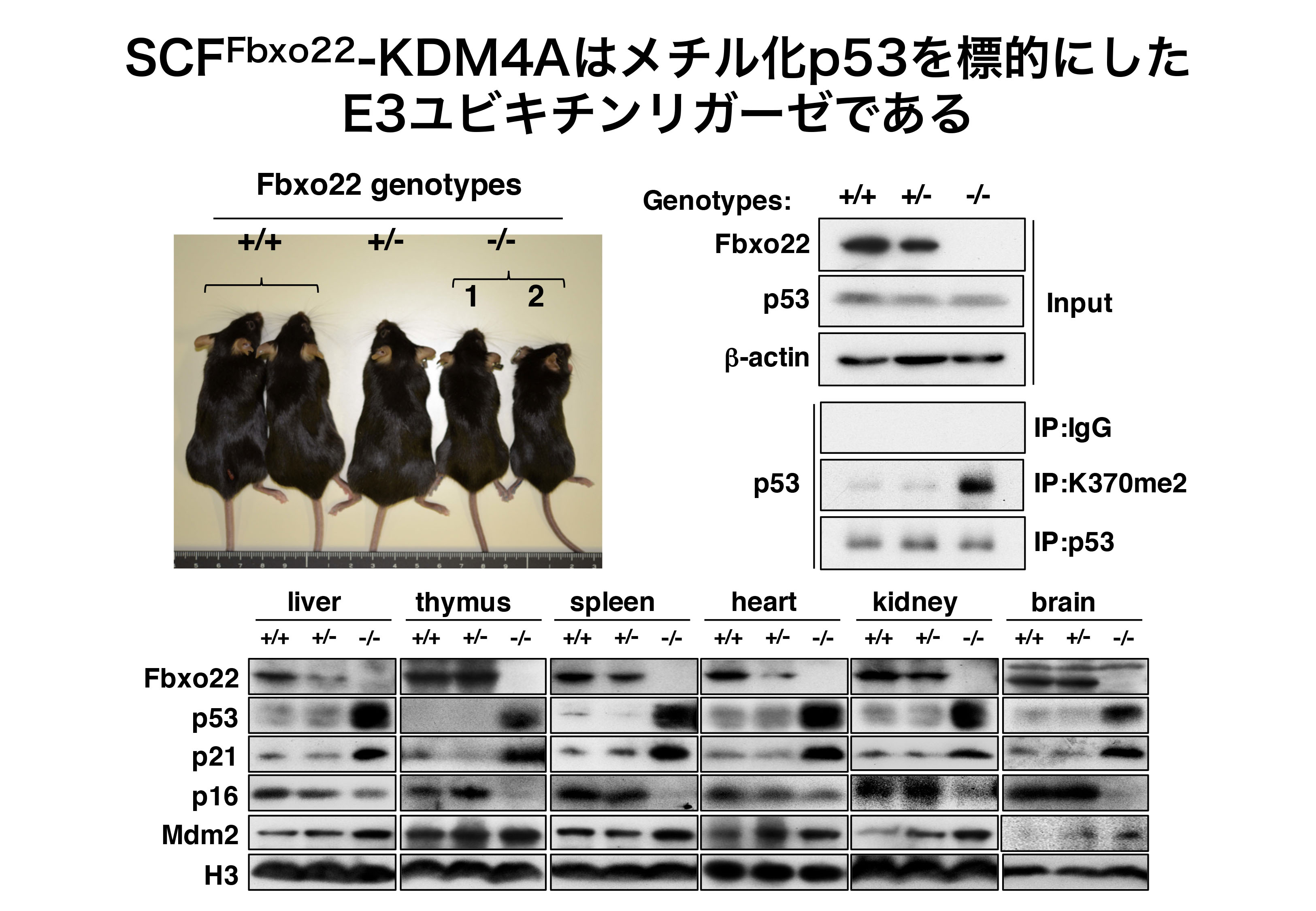
面白いことに、Fbxo22ノックアウトマウスは、様々な臓器・組織においてメチル化p53の蓄積が起こり、細胞の増殖が抑制されるため、 正常やヘテロマウスに比較して約半分の大きさであった(図1)。
またこれらのマウス由来の細胞を用いた研究から、老化細胞におけるFbxo22の特異的な発現は、老化後期にp53を分解して 不活性化することで、SASPやp16発現誘導に必須に役割を果たしていることが明らかとなった(図2)。