1. 細胞老化誘導機構の解明
Hayflickらにより,ヒトの正常な体細胞は培養皿で継代をくり返すと最終的に不可逆的な細胞増殖の停止をひき起こすことがはじめて示された。 のちの解析により,幹細胞を除く多くのヒトの正常な細胞にはテロメアの短小化に起因する細胞分裂の回数の限界(分裂寿命)のあることがわかり, 分裂寿命により生じた細胞老化はとくに“複製老化”と定義づけられた。
一方,細胞老化に関する研究が進むにつれ,Ras遺伝子などのがん遺伝子の活性化,酸化ストレス,DNA損傷などによっても細胞老化が誘導されること も明らかになり,これらは“ストレス性の細胞老化”などとよばれている.細胞老化のもっとも重要かつ共通の特徴のひとつは不可逆的な 細胞増殖の停止であり,その特徴から,細胞死を誘導するアポトーシスとならんで,もっとも重要な抗腫瘍化機構のひとつであると考えられている。
また,近年の研究により,老化細胞をマウス個体から人工的に除去すると加齢性変化が遅れ、寿命が延長することが示されたことから, 細胞老化は加齢にともなう病態にも深く関与することが明らかになりつつある。
我々は、老化細胞が細胞周期のどのステージにおいて不可逆的に細胞増殖を停止するかを検討するため,FUCCI(fluorescent ubiquitination-based cell cycle indicator)系とよばれる細胞周期の動的な変化を生細胞イメージングにより観察する方法を応用することにより,細胞老化の過程における細胞周期の 変化を1細胞レベルで解析した.
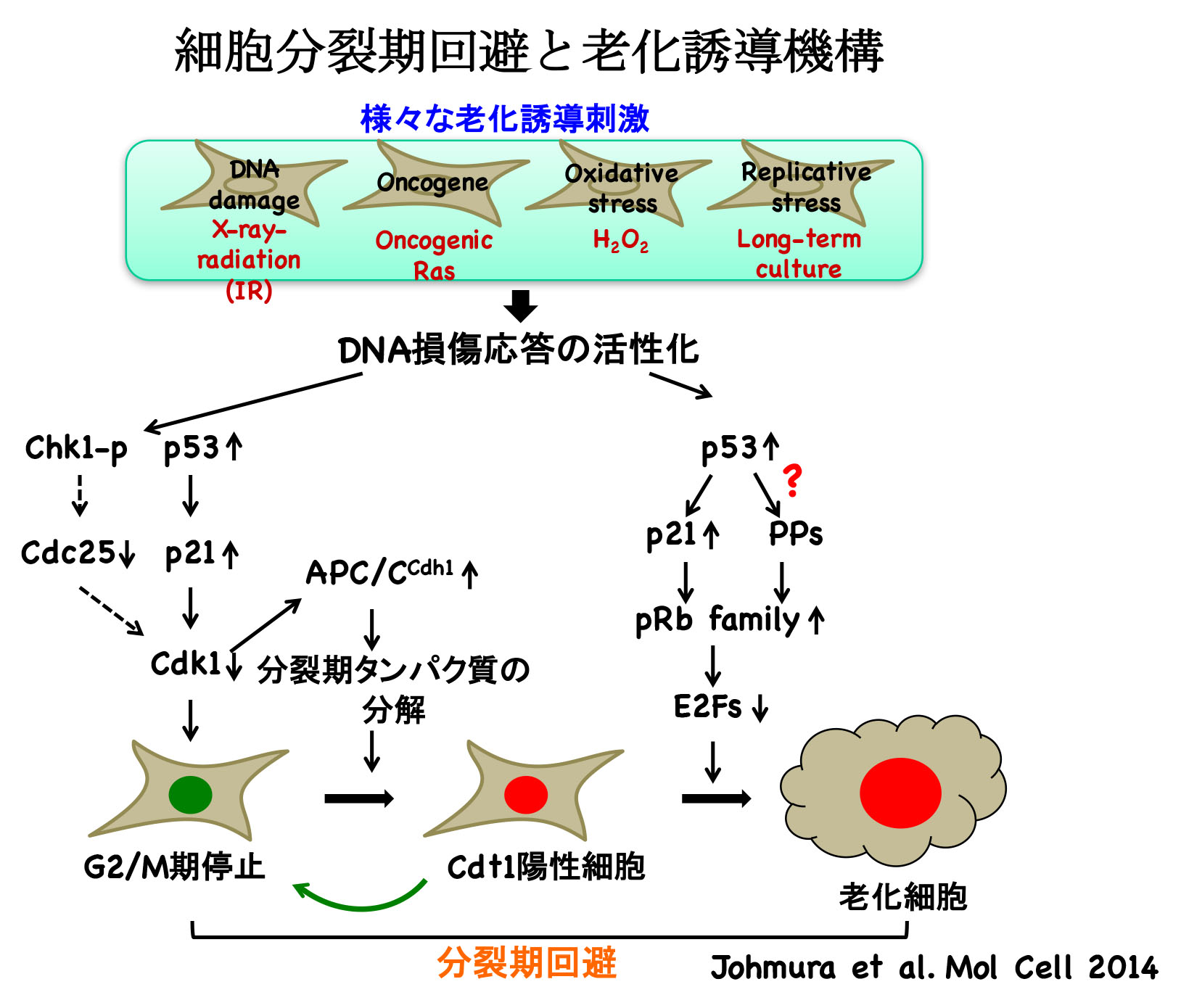
その結果,がん遺伝子のストレスを含めたすべての細胞老化の過程において,多くの細胞がG2期における一時的な細胞周期の停止ののち, 通常は起こるべき細胞分裂をすることなくG1期に進行する,いわゆる,分裂期の回避を生じて4倍体のG1期細胞となり,不可逆的に細胞周期を 停止することを見い出した.また,老化の過程における分裂制御タンパク質の発現の変化を解析したところ,解析したすべてのタンパク質の発現が分裂期の回避 のタイミングにあわせていちじるしく低下していることがわかった.この結果は,分裂制御タンパク質の発現の顕著な低下が分裂期の回避の引き金になっていることを示唆した。
さらに,DNA損傷により細胞老化を誘導した細胞を蛍光セルソーターにより2倍体細胞と4倍体細胞とに分画したのち, 細胞老化のマーカーであるCDK4およびCDK6の阻害タンパク質であるp16の発現の変化の解析,および,SA-β-gal(senescence-associated β-galactosidase)染色を行った。その結果,4倍体細胞はp16の発現の顕著な上昇を示すこと,そのほとんどがSA-β-gal陽性細胞で あることがわかった。
これらの結果から,老化細胞は分裂期の回避により生じた四倍体のG1期細胞であると考えられた.
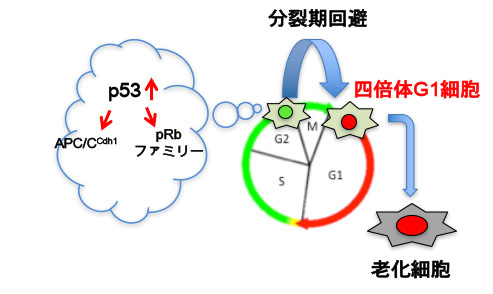
分裂期の回避が誘導される分子機構および細胞老化の誘導における役割について調べた.がん抑制遺伝子の産物 であるp53は細胞老化の誘導の鍵となるタンパク質であることが知られていることから,RNAi法を用いて, p53遺伝子の発現の抑制をテトラサイクリンにより誘導できる細胞株を樹立し,FUCCI系により生細胞イメージング 解析を行った.その結果,対照となる細胞と比較して,p53の発現を抑制した細胞の多くにおいては老化を誘導する 種々の刺激に対し分裂期の回避は起こらないことがわかった.また,このとき,SA-β-gal陽性細胞の顕著な減少も 認められた。
以上より,p53は分裂期の回避および細胞老化の誘導に必須であることが示された.
以前の報告より,p53の単純な活性化だけでは細胞老化を誘導できないと考えられていた.そこで、細胞周期に
特異的かつ一過的にp53の発現を制御できる系を構築するため,レンチウイルスベクターを用いて、p53遺伝子の
発現をテトラサイクリンにより誘導できる細胞株を樹立した.この細胞をCDK1の阻 害剤によりG2期に同調させたのち、ドキシサイクリンで48時間処理することによりp53を一過的に発現させたところ, 多くの細胞において分裂期の回避および不可逆的な細胞増殖の停止が認められた。
一方,血清飢餓やDNA複製阻害剤などによりG0期やG1期/S期に同調した細胞に一過的にp53を発現させても細胞老化の誘導は認められなかった。
また,p53の分解においてはたらくユビキチンリガーゼとして知られるMdm2の阻害剤を用いてp53の発現を一過的に上昇させた場合にも同様の結果が得られた。
以上の結果より,G2期におけるp53の活性化が分裂期の回避および細胞老化の誘導に必要かつ十分である ことが明らかになった.さらに解析を進めたところ、p53はAPC/Ccdh1およびpRbファミリーを協調的に活性化することにより分裂期の回避および 細胞老化を誘導する可能性が考えられた.この可能性を検討するため,Cdh1およびpRbの構成的な活性化変異体それぞれ単独の発現,あるいは,その両方 の発現をテトラサイクリンにより誘導できる細胞株を樹立して解析した.その結果,p53を過剰に発現する細胞と同様に,Cdh1およびpRbの発現をG2期において 一過的に誘導した場合のみ,分裂期の回避および細胞老化の誘導が認められた。
分裂期の回避がヒトの生体においても細胞老化の誘導において重要な役割をはたしているかどうかを検討した.ヒトの母斑細胞(ほくろ)はRas遺伝子の下流 ではたらくリン酸化酵素BRAFの変異により生じる老化細胞であることが報告されている.そこで,母斑の組織切片における細胞のDNA含量を測定するため DAPI染色を行った.その結果,正常な上皮細胞あるいは血管内皮細胞と比較して,母斑細胞では約2倍のDNA含量を示すことがわかった.さらに,G2期 /M期のマーカーであるサイクリンB1や細胞増殖のマーカーであるKi67による組織免疫染色を行ったところ,母斑細胞においてはどちらの発現も認められ なかった.これらの結果から,生体に存在する老化細胞も4倍体のG1期細胞であることが示唆された.
現在G1期の細胞において、倍の染色体数がどのようなシグナルを、どのような分子機構で発信して増殖停止を誘導しているのかについてさらに解析を進めている。