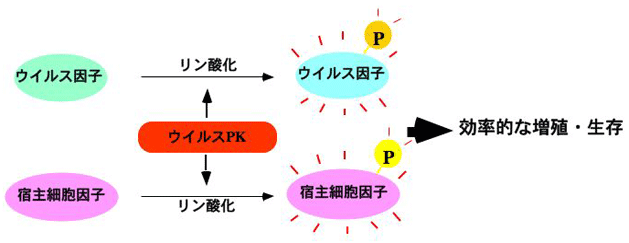
ウイルスPKの機能発現機構解明の第1ステップは、標的基質を同定することにある。しかし、ウイルスPKに対するウイルス側の標的因子が次々に同定されたのに対し、宿主細胞標的因子に関わる知見は皆無であった。我々は、HSVがコードするPKであるUL13が宿主翻訳因子EF-1 deltaをリン酸化することを明らかにした。本知見は、ウイルス特異PKの宿主細胞標的因子を、ウイルス学ではじめて同定したものとなった。
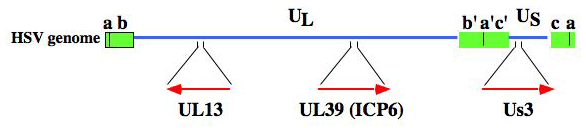
ヘルペスウイルスがコードするウイルスタンパク質は、(i) 全てのヘルペスウイルスに共通に保存されているもの、(ii) ヘルペスウイルスの各亜科でのみ保存されているもの、(iii) 各ヘルペスウイルスに特異的なものに分類される。UL13は全てのヘルペスウイルスにおいて保存されており、ヘルペスウイルス感染において普遍的な役割を果たしていることが示唆されていた。実際我々は、EF-1 deltaのリン酸化は、全ての亜科のヘルペスウイルス感染において共通に引き起こされ、そのリン酸化にはヘルペスウイルスで保存されているPK (CHPK: conserved herpesvirus protein kinase)が関与していることを明らかにした。つまり、CHPKの普遍的な役割の1つがEF-1 deltaのリン酸化であることが明らかになった。
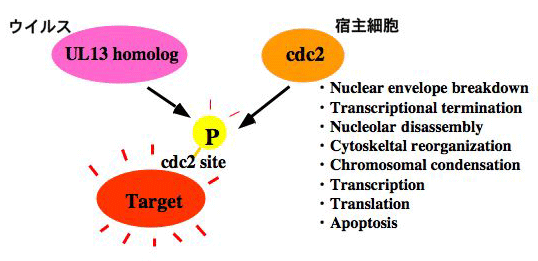
UL13によるEF-1 deltaのリン酸化の生物学的意義を解明するために、リン酸化部位の同定を試みた。その結果、非感染細胞および感染細胞の双方におけるEF-1のリン酸化部位が133番目のセリンであることが明らかとなった。この知見は、非感染細胞における宿主細胞PKと感染細胞におけるHSV UL13がEF-1 deltaの同一アミノ酸残基(Ser-133)をリン酸化する、つまり、機能的にHSV UL13と相同性を示す宿主細胞PKの存在を示唆していた。そこで、この宿主細胞PKを同定したところ、宿主細胞PK cdc2であった。興味深いことに、UL13とcdc2がEF-1 deltaの同一部位をリン酸化するという事象は、UL13-EF-1 deltaの組み合わせに限られた事象ではなかった。つまり、ヘルペスウイルスで保存されているCHPKsと宿主細胞PK cdc2は、標的因子の同一アミノ酸残基をリン酸化することが明らかになった。このように、我々はCHPKの存在意義が宿主PK cdc2の模倣であることを明らかにした。Cdc2は、転写、翻訳、細胞骨格、核膜、クロマチンといった様々な細胞機構を制御している。Cdc2様の活性をもつPKをウイルスが保持することは、「宿主細胞機構をのっとる」というウイルスの目的遂行のためには、大きなメリットであると考えられる。