
浸透圧ストレスに対する細胞応答の動的制御機構を解明
浸透圧ストレスに対する細胞応答の動的制御機構を解明
生体は常に環境の変化に曝されています。生命を維持するには変化に対する適切な応答が欠かせません。例えば、高浸透圧ストレスに対する適応も、すべての生物に共通した基礎的な生命現象です。ヒトの体液では、浸透圧が正常値から10%程度上下しただけで生命にかかわるほどです。これほど重大なものでありながら、細胞がどのようにして浸透圧を感知し、どのようにして適応反応を引き起こすのかは、よくわかっていません。この論文は、酵母細胞をモデル生物として使い、浸透圧ストレス応答の精緻な制御機構の一端を解明したものです。
酵母が高浸透圧ストレスに曝されると、大量にグリセリンを合成することによって細胞内外の浸透圧差を解消します。この応答を制御する機構の中心にあるのが、Ste20-Ste11-Pbs2-Hog1の4種のキナーゼよりなるHog1 MAPK経路です。細胞膜中の浸透圧センサーHkr1とMsb2とが浸透圧を感知すると、Cdc42 Gタンパク質がGTP結合型に変換され、Ste20に結合して活性化します。Cdc42-GTPは膜に結合しているので、活性化したSte20も膜に局在します。したがって、Ste11がSte20によって活性化されるためには、通常は細胞質にいるSte11を膜に局在化する必要があります。この反応をつかさどるのがアダプタータンパク質Ste50で、Ste11と膜タンパク質Opy2とに同時に結合することで、Ste11を膜に局在させます。
このOpy2にはSte50と結合する領域が三つ(CR-A、CR-B、CR-D)あることがわかりました。CR-AとCR-Dは恒常的にSte50と結合できますが、CR-Bは、グルコース存在下で活性化するキナーゼ(Yck1/Yck2)にリン酸化されてはじめてSte50に結合できるようになります(図A)。したがって、環境にグルコースが存在すると、より多くのSte50がOpy2に結合し、浸透圧に対する応答性が高くなると考えられます。天然に生育する酵母にとっての浸透圧ストレスは熟れた果実の糖分に起因するものであることが多いので、グルコース増加は浸透圧上昇の先触れになります。両者が浸透圧応答を協同的に制御するのはきわめて合理的なことと言えるでしょう。
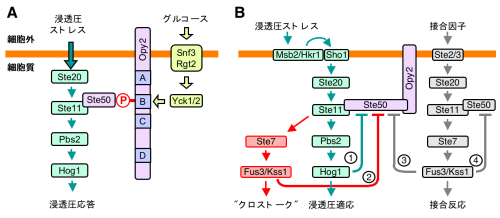
一方、浸透圧により活性化したHog1キナーゼや接合因子により活性化したFus3キナーゼがSte50をリン酸化し、その結果Ste50がOpy2から解離することもわかりました(図B)。この反応には、Hog1キナーゼの過剰な活性化を防ぎ、また接合反応進行中は浸透圧応答を抑制する、という機能があります。
結論として、リン酸化反応によるSte50とOpy2との結合の増強あるいは減弱が、浸透圧ストレス由来の情報とグルコースや接合因子由来の情報とを統合していることが明らかになりました。