研究内容
当研究室では、より効果的なワクチン開発に役立てることを目的として、生体によるウイルス認識機構、インフルエンザウイルス感染モデルを用いた粘膜免疫制御機構と腸内細菌によるウイルス特異的免疫応答の制御機構について研究を行っております。
・研究内容
・発表論文解説
研究内容
1.自然免疫系によるウイルス認識機構
⇒生体が目には見えないウイルスをどう異物として認識しているのか?
・Moriyama et al. iScience. 2020 [Link]
・Moriyama et al. Nat Commun. 2019 [Link] [プレスリリース]
・Chen et al. Front Microbiol. 2019 [Link]
・Ichinohe et al. PNAS. 2013 [Link] [プレスリリース]
・Ito et al. PLoS Pathog. 2012 [Link]
・Ichinohe et al. Nat Immunol. 2010 [Link]
2.腸内細菌叢によるウイルス性肺炎の重症化抑制機構
⇒腸内細菌叢がウイルス性肺炎の重症化の抑制にどう役立っているのか?
・Nagai et al. Nat Commun. 2023 [Link] [プレスリリース]
・Moriyama et al. PNAS. 2019 [Link] [プレスリリース]
・Ichinohe et al. PNAS. 2011 [Link]
3.ウイルスの宿主免疫回避機構
⇒ウイルスは宿主の免疫をどう回避しているのか?
・Moriyama et al. J Virol. 2018 [Link]
・Moriyama et al. J Virol. 2016 [Link]
・Komune a et al. J Virol. 2011 [Link
4.ウイルスの病原性発現機構
⇒インフルエンザウイルスや新型コロナウイルスがヒトや動物を重症化させるメカニズムは何か?
⇒武漢株、デルタ株、オミクロン株で重症度に違いがあるのはなぜか?
・Kobayashi et al. PLoS Pathog. 2024 [Link]
5.ウイルス感染による重症化メカニズムの解明
⇒なぜ高齢者ではインフルエンザウイルスや新型コロナウイルスに感染すると重症化しやすいのか?
・Nagai et al. Nat Commun. 2023 [Link] [プレスリリース]
6.ウイルスと宿主の共生メカニズム
⇒水きん類(アヒルやカモなどの水鳥)はなぜインフルエンザウイルスと共生できるのか?
⇒新型コロナウイルスに感染しても無症状のひとがいるのはなぜか?
・Nagai et al. Nat Commun. 2023 [Link] [プレスリリース]
7.インフルエンザウイルスや新型コロナウイルスに対する経鼻ワクチンの開発
⇒鼻に噴霧してウイルスの感染そのものを阻止する次世代ワクチンの研究
・Kobayashi et al. NPJ Vaccines. 2025 [Link]
・Nagai et al. mBio. 2021 [Link] [プレスリリース]
・Ichinohe et al. PNAS. 2011 [Link]
・Ichinohe et al. J Exp Med. 2009 [Link]
8.±1℃の生体防御学
⇒発熱の意義とは何か?1℃の違いがウイルス増殖に与える影響は?
・Kobayashi et al. 論文投稿中
・Nagai et al. Nat Commun. 2023 [Link] [プレスリリース]
・Moriyama et al. PNAS. 2019 [Link] [プレスリリース]
参考:論文引用回数(Ichinohe et al. PNAS 2011)

発表論文解説
| Kobayashi M, Kobayashi N, Deguchi K, Omori S, Ichinohe T. SARS-CoV-2 infection primes cross-protective respiratory IgA in a MyD88- and MAVS-dependent manner. NPJ Vaccines. 2025 Feb 27;10(1):40. |
新型コロナウイルス(SARS-CoV-2)は、mRNAワクチンにより誘導された血液中のIgG抗体から逃れるための戦略として、スパイクタンパク質に変異を導入して進化を続けています。インフルエンザウイルスを用いたこれまでの研究では、呼吸器粘膜に誘導されるウイルス特異的なIgA抗体は、変異株に対する感染防御効果に優れていることが分かっていました。一方、SARS-CoV-2特異的なIgA抗体の変異株に対する感染防御効果についてはよく分かっていませんでした。
呼吸器粘膜に誘導されたSARS-CoV-2特異的なIgA抗体の変異株に対する感染防御効果を調べるため、マウスにSARS-CoV-2オミクロンBA.1株を10 pfu(plaque-forming unit)感染させ、感染1〜5週間後までの血清と肺胞洗浄液中を回収し、スパイク特異的なIgGおよびIgA抗体を解析しました(Figure 1a)。オミクロンBA.1株の感染により、血液中および呼吸器粘膜にスパイク特異的なIgG抗体が誘導されました(Figure 1b, c)。一方、オミクロンBA.1株の感染では、呼吸器粘膜にスパイクIgA抗体が有意に誘導されていないことが分かりました(Figure 1d)。そこでオミクロンBA.1株を感染させてから5週間後に、スパイクタンパク質5μgを経鼻投与(追加免疫)すると、呼吸器粘膜にスパイク特異的なIgA抗体が有意に誘導されることが分かりました(Figure 1e-g)。
呼吸器粘膜に誘導されたスパイク特異的なIgA抗体の変異株に対する感染防御効果を調べるため、マウスにSARS-CoV-2オミクロンBA.1株を10 pfu感染させ、5週間後にスパイクタンパク質5μgを筋注または経鼻で追加免疫し、その2週間後に致死量の新型コロナウイルスガンマ株(1×106 pfu)を感染させました(Figure 2a)。感染2〜3日後の呼吸器粘膜に検出される感染性ウイルス量を解析すると、筋注で追加免疫したグループ(呼吸器粘膜にスパイク特異的なIgA抗体を持たない)は、1mlあたり106〜107 pfuの感染性ウイルスが検出されましたが、経鼻で追加免疫したグループ(呼吸器粘膜にスパイク特異的なIgA抗体がある)では、感染性ウイルスが検出限界以下でした(Figure 2e)。筋注で追加免疫したグループでは、ガンマ株感染後に有意な体重減少が認められ3割のマウスが死亡したのに対して、経鼻で追加免疫をして呼吸器粘膜にスパイク特異的なIgA抗体を誘導したグループでは、ガンマ株感染による体重減少は認められず、すべてのマウスが生存しました(Figure 2f, g)。
最近の研究では、細胞傷害性T細胞(CD8T細胞)が新型コロナウイルスの変異株に対する感染防御効果に重要であることが報告されています(参考文献14, 15)。新型コロナウイルス変異株の感染防御効果におけるCD8T細胞の役割を調べるため、マウスにSARS-CoV-2オミクロンBA.1株を10 pfu感染させ、5週間後にスパイクタンパク質5μgを経鼻で追加免疫したあと、CD8に対する抗体を経鼻および静脈投与して、マウスの体内からCD8T細胞を枯渇(depletion)させました(Figure 3a)。CD8に対する抗体を経鼻および静脈投与により、マウスの体内からCD8T細胞が消えていることを確認したあと(Supplementary Figure 7)、致死量の新型コロナウイルスガンマ株(1×106 pfu)を感染させ、呼吸器粘膜の感染性ウイルス量や体重変化、生存率を解析しましたが、生体内のCD8T細胞を枯渇させても、気道粘膜のスパイク特異的なIgA抗体があれば、新型コロナウイルス変異株(ガンマ株)の感染を完全にブロックできることが分かりました(Figure 3c-e)。
以上のことから、スパイクタンパク質の経鼻的な追加免疫により誘導された気道粘膜のスパイク特異的なIgA抗体は、新型コロナウイルス変異株に対する感染防御に重要であることが明らかとなりました。最後にこのスパイク特異的なIgA抗体の誘導に必要な自然免疫シグナルを解析したところ、RNAウイルス認識経路に必要なMyD88やMAVSが重要な役割を果たしていることが明らかとなりました(Figure 4, Figure 5)。本研究成果は、気道粘膜のIgA抗体が新型コロナウイルス変異株に対する感染防御に重要であることを示しただけでなく、新型コロナウイルス変異株に対しても有効な経鼻ワクチンの開発に繋がる重要な知見であると言えます。

Q1.新型コロナウイルスはマウスに感染するのですか?新型コロナウイルスはマウスには感染しないので、新型コロナの研究にはハムスターなどが用いられていると思っていたのですが。
A1.新型コロナウイルスはマウスに感染しません。そこで我々は新型コロナウイルスをマウスで継代(マウスに繰り返し感染させること)し、マウスにも感染する「マウス馴化新型コロナウイルス」を樹立しました(Supplementary Figure 3)。新型コロナの研究に、ハムスターが用いられることもありますが、ハムスターではワクチン接種後の免疫学的な解析が難しい(IgG応答しか測れない)ので、本研究ではマウス馴化新型コロナウイルスを使い、血中のスパイク特異的なIgG抗体だけでなく、気道粘膜のスパイク特異的なIgA抗体を測定しています。世の中にはさまざまな遺伝子欠損マウスが揃っているので、マウスを使った新型コロナウイルス研究は、ワクチン開発研究だけでなくCOVID-19重症化メカニズムの研究(例えばどういう遺伝子欠損がCOVID-19の重症化を引き起こすのかなど)を行うことができるというメリットもあると思います。
Q2.授業で「インフルエンザウイルスに感染すると、気道粘膜にウイルス特異的なIgA抗体が誘導される」と習いました。今回、マウス馴化新型コロナウイルスBA.1を感染させても、気道粘膜に十分なIgA抗体が誘導されなかったということですが、これはなぜですか?
A2.マウスの系統(stain)の差があるかもしれません。Balb/cマウスにマウス馴化新型コロナウイルスBA.1株を感染させると、5週間後には気道粘膜にスパイク特異的なIgA抗体が誘導されることを確認しています(Supplementary Figure 5)。本研究でC57BL6/Jマウスを使ったのは、多くの遺伝子欠損マウスの遺伝的背景(系統)が、このC57BL6/Jマウスと同じだから(野生型マウスと遺伝子欠損マウスの系統を揃えるため)です。
新型コロナウイルス感染症(COVID-19)は、社会的および経済的に深刻なダメージを与えました。ウイルス感染によって誘発されるサイトカインストームがCOVID-19の重症化と関連しているという証拠が増えている一方で、炎症性サイトカインのCOVID-19重症化における役割は未だ不明です。本研究では、致死量のマウス馴化新型コロナウイルス(デルタ株)をマウスへ感染させることにより、MyD88およびI型インターフェロン受容体(IFNAR1)シグナルを介して、炎症性サイトカインの産生が増幅されることを見出しました。また炎症性サイトカインのTNF-αは肺胞マクロファージに作用するとCXCL1(好中球の遊走に必要なケモカイン)の産生を増幅させ、感染局所に好中球を浸潤させて、ウイルス感染後の重症度を悪化させることを明らかにしました。さらに、ウイルス感染後にTNF-α分泌阻害剤や抗TNF-α抗体を静脈内投与することにより、マウスおよびハムスターの重症度を部分的に軽減させることができることを示しました。本研究成果は、COVID-19の重症化メカニズムの一端を明らかにしたものであり、COVID-19に限らず他の呼吸器ウイルス感染症および関連疾患の新しい治療戦略に貢献する重要な知見であると言えます。

図.COVID-19重症化メカニズム
致死量の新型コロナウイルス感染を感染させると、MyD88/IFNAR1依存的に炎症性サイトカインが増幅する。TNF-αはマクロファージに作用するとCXCL-1の産生を促進させる。これにより感染局所に好中球が浸潤して過剰な炎症反応を引き起こし、肺炎が悪化する。
Q1.インターフェロンはウイルスの感染防御に必要ではないのですか?
A1.必要です。実際にマウス馴化SARS-CoV-2祖先株やデルタ株の感染と同時にマウスの組換えIFN-α/βを経鼻投与すると、ウイルスの感染を完全にブロックすることができました(Figure 6A-C)。一方、マウスを重症化させないマウス馴化SARS-CoV-2祖先株を感染させて2〜4日後にIFN-α/βを投与するとマウスが重症化することが分かりました(Figure 6D-F)。このことからインターフェロン応答のタイミングにより、ウイルス感染防御に働く場合と症状を悪化させてしまう場合があることが示唆されました。
Q2.COVID-19重症患者では、インターフェロン応答関連遺伝子に欠損が認められる場合やI型インターフェロンに対する自己抗体があることが確認されています(参考文献27-29)。ヒトで確認されているこれらの事実と今回の結果(MyD88欠損やIFNAR1欠損マウスはマウス馴化SARS-CoV-2デルタ株感染後に重症化しにくいこと)は矛盾しませんか?
A2.これは感染するときのウイルスの量(dose)による違いではないかと考えています。A1.でも述べましたように、感染と同時のインターフェロン応答はウイルスの感染防御に必要です。COVID-19重症患者で見られているような遺伝子欠損やインターフェロンに対する自己抗体は、宿主の抗ウイルス自然免疫応答を減弱させ、少量のウイルスであっても宿主の抗ウイルス自然免疫応答の壁を突破してしまい、結果として体内で大量のウイルスが増殖し、重症化を引き起こしている可能性があります。また本研究で用いたような致死量(1×10^5 pfu)のウイルスを感染させた場合にも、ウイルスが宿主の抗ウイルス自然免疫応答の壁を突破(インターフェロン応答が感染初期のウイルス複製を抑制することに失敗)することになり、過剰な炎症や肺組織ダメージを引き起こしている可能性があります。本研究では、MyD88欠損マウスに非致死量(1×10^4 pfu)のマウス馴化SARS-CoV-2デルタ株を感染さる実験も行いましたが、野生型マウスと比較して体重変化や生存率に違いは認められませんでした(Supplementary Figure 8)。
Q3.好中球が肺組織に遊走するとなぜ症状を悪化させてしまうことになるのですか?
A3.感染局所のケモカイン(CXCL1)を目印に好中球が遊走してきます。感染局所に遊走してきた好中球は異物を認識すると核からDNAを放出し、好中球細胞外トラップ(NETs; neutrophil extracellular traps)を作ります。地引網のときに使うような大きなネットです。このNETsはウイルスや細菌などの微生物の補足に役立つ一方、宿主にとって細胞外のDNA(NETs)は異物となるため、自然免疫を活性化させて余計な炎症応答を引き起こしてしまうというリスクがあるからです。好中球がCOVID-19の重症化と関連しているという報告がいくつかありますので参考にしてみてください(参考文献10-14)。
これまで外気温や体温がウイルスに感染した場合の重症度に与える影響についてはほとんど分かっていませんでした。外気温や体温がウイルス感染後の重症度に与える影響を解析するため、さまざまな温度条件で飼育したマウスにインフルエンザウイルスを感染させた場合の重症度を解析しました。すると36℃条件下で飼育したマウスでは体温が38℃を超えるようになり、インフルエンザウイルスのみならずSARS-CoV-2の感染に対しても高い抵抗力を獲得することが分かりました。22℃で飼育したマウス(体温は37℃前後)と36℃条件下で飼育して体温が38℃を越えたマウスの血清と盲腸内容物のメタボローム解析を行ったところ、体温が38℃を越えたマウスの体内では胆汁酸レベルが有意に増加しており、特に盲腸内容物中では二次胆汁酸量が有意に増加していることを見出しました。また22℃で飼育したマウスやハムスターにデオキシコール酸(DCA)やウルソデオキシコール酸(UDCA)などの二次胆汁酸を与えると、インフルエンザウイルスやSARS-CoV-2感染後の生存率が有意に改善することを明らかにしました。さらにCOVID-19患者から採取した血液サンプルを解析したところ、胆汁酸レベルが軽症患者グループと比較して中等症I/II患者グループで有意に低下していることが明らかとなりました。このことはヒトにおいてもCOVID-19の重症度と胆汁酸レベルに逆相関関係があることを示しています。本研究成果は、体温が38℃以上に上昇することにより腸内細菌叢が活性化し、二次胆汁酸産生を介してウイルス感染後の重症化予防に役立っていることを分子レベルで明らかにした世界初の成果であり、高齢者がインフルエンザやCOVID-19で重症化しやすくなるメカニズムの解明や、宿主とウイルスの共生メカニズムの解明、胆汁酸受容体を標的としたウイルス性肺炎の重症化を抑える新しい治療薬の開発などに繋がることが期待されます。
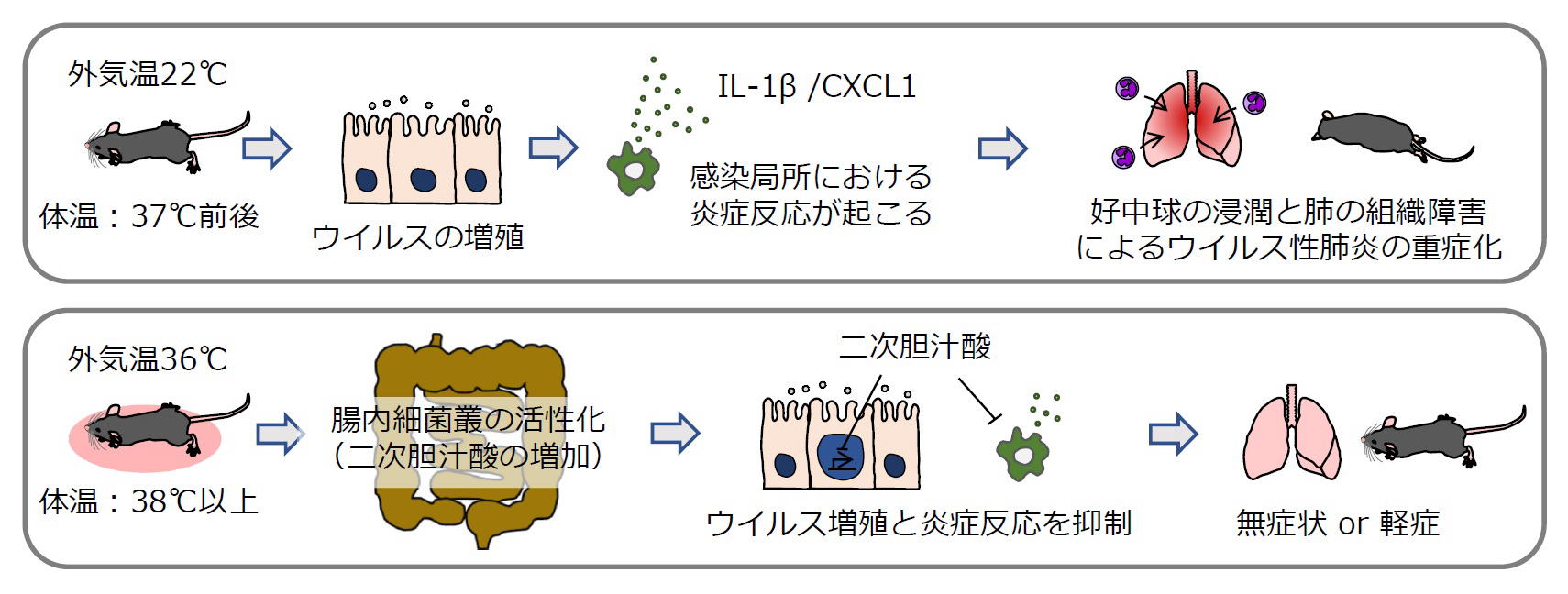
図.本研究成果のまとめ
上段:22℃で飼育したマウス(体温は37℃前後)にインフルエンザウイルスを感染させるとウイルスが増殖し、マクロファージなどからサイトカインやケモカイン(IL-1β、CXCL1など)が産生され、感染局所の炎症が起こる。感染局所の炎症を目印に肺へ遊走してきた好中球が炎症を増大させて肺の組織障害を引き起こし、ウイルス性肺炎が重症化する。
下段:36℃で飼育したマウス(体温は38℃以上)では腸内細菌叢が活性化することにより、血中の二次胆汁酸レベルが増加する。二次胆汁酸はウイルスの増殖および炎症を誘発するサイトカイン、ケモカインの産生を抑制する。その結果、肺における過剰な炎症が起きることなく、ウイルスに感染したマウスは無症状や軽症となる。
・プレスリリース
| Moriyama M, Nagai M, Maruzuru Y, Koshiba T, Kawaguchi Y, Ichinohe T. Influenza virus-induced oxidized DNA activates inflammasomes. iScience. 2020 Jun 14;23(7):101270. |
インフルエンザウイルスのM2タンパク質やPB1-F2タンパク質は炎症反応に関わることが知られています(Ichinohe et al. Nat Immunol. 2010, McAuley et al. PLoS Pathog. 2013)。しかし、これらのウイルスタンパク質がどのように炎症反応を引き起こしているのか、その詳細なメカニズムは不明でした。今回、インフルエンザウイルスを感染させたマクロファージでは、核やミトコンドリア由来のDNAが細胞質中やマクロファージ細胞外トラップと呼ばれるネット状の構造物中に多く検出できることを見出しました。これらのDNAには酸化DNAが含まれており、インフルエンザウイルスの複製に必須のM2タンパク質が酸化DNAの放出を引き起こしていることを明らかにしました。またミトコンドリア内に局在するPB1-F2タンパク質は、ウイルスRNA存在下で酸化DNAの放出を誘導しました。核やミトコンドリア由来と考えられるこれらの酸化DNAは、NLRP3やAIM2インフラマソーム依存的なIL-1βの産生を誘導しており、これがウイルス感染局所の炎症応答に関わっていることが示唆されました。M2タンパク質のようなイオンチャネル活性を持つウイルスタンパク質は新型コロナウイルスを含む他のウイルスにも認められることから、本研究成果はインフルエンザウイルスだけでなく、新型コロナウイルスなどの強い炎症反応を引き起こすウイルスの病原性発現機構の解明にも繋がると期待されます。
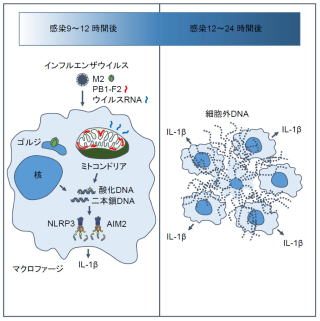
図.本研究成果のまとめ
インフルエンザウイルスがマクロファージに感染すると、ウイルスのM2タンパク質の作用により、酸化DNAが細胞質中へ放出する。ミトコンドリアに局在するPB1-F2タンパク質は、ウイルスRNA存在下で酸化DNAを放出させる。酸化DNAを含む核やミトコンドリア由来の二本鎖DNAはNLRP3やAIM2インフラマソーム依存的なIL-1βの分泌を引き起こす。細胞質中のDNA量はインフルエンザウイルス感染8〜9時間後にピークとなる一方、ウイルス感染12時間後をピークに細胞外にも二本鎖DNAが検出できるようになる。さらにウイルス感染24時間後にはマクロファージ細胞外トラップと呼ばれるネット状の構造物中にDNAを検出できるようになる。この細胞外DNAは周囲のマクロファージに作用してIL-1βの産生を増大させていると考えられる。
・プレスリリース
| Moriyama M, Koshiba T, Ichinohe T. Influenza A virus M2 protein triggers mitochondrial DNA-mediated antiviral immune responses. Nat Commun. 2019 Oct 11;10(1):4624. |
DNAウイルスが細胞に感染すると細胞内のDNAセンサーがウイルスDNAを認識してインターフェロン応答を誘導することが知られています。最近の研究では、一部のRNAウイルスが細胞に感染した場合でも細胞質中にミトコンドリアDNAが放出されて、細胞内のDNAセンサーを介してインターフェロン応答が誘導されることが報告されております。しかしRNAウイルスがどのようにミトコンドリアDNAを放出させるのかは不明でした。今回、インフルエンザウイルスの複製に必須のM2タンパク質がミトコンドリアDNAの放出を引き起こしていることを突き止めました。インフルエンザウイルスの感染によって細胞質中に放出されたミトコンドリアDNAは、細胞内のDNAセンサーであるcGASやDDX41とそのアダプタータンパク質であるSTINGを介してインターフェロンβを誘導していました。さらにSTINGを欠損したマウスでは、インフルエンザウイルス感染5日目の肺のウイルス量が、野生型マウスと比較して有意に増加していたことから、このSTING依存的なインターフェロン応答が生体内でインフルエンザウイルスの増殖を抑制するのに必須であることも明らかとなりました。さらにインフルエンザウイルスのNS1タンパク質はミトコンドリアDNAと結合することにより、細胞内のDNAセンサーからミトコンドリアDNAが検出されることを逃れていることも明らかとなりました。本研究成果は、インフルエンザウイルスワクチンの効果を高めるアジュバントの開発や、インフルエンザウイルスが効率よく増殖するメカニズムの解明、インフルエンザウイルスの病原性発現機構の解明に繋がると期待されます。
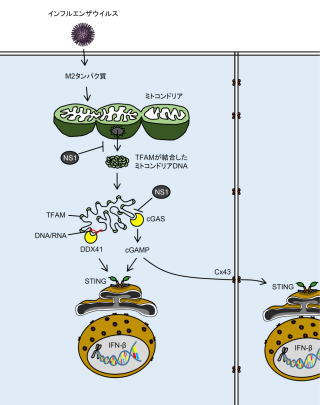
図.本研究成果のまとめ
インフルエンザウイルスが細胞に感染すると、ウイルスのM2タンパク質の作用により、ミトコンドリアにダメージを与えてミトコンドリアDNAが細胞質中へ放出される。細胞質中へ放出されたミトコンドリアDNAはTFAMが結合した状態であると考えられるものの、細胞内のDNAセンサーであるcGASやDDX41は細胞質中ミトコンドリアDNAまたはDNA/RNAハイブリッド鎖を認識したあと、その下流のアダプタータンパク質であるSTINGを介してインターフェロンβを誘導する。cGASにより合成されたcGAMPはギャップ結合を構成するconnexin 43(Cx43)を介して隣接する細胞へ拡散され、STING依存的なインターフェロン応答を増幅させている。インフルエンザウイルスのNS1タンパク質は、ミトコンドリアDNAの放出そのものを抑制しているか、細胞質中に放出されたミトコンドリアDNAに結合することにより、ミトコンドリアDNAが宿主のDNAセンサーから認識されるのを逃れている。
・プレスリリース
・東大新聞オンライン


(2019年10月22日掲載。東京大学新聞社が記事転載を許可しています)
| Moriyama M, Ichinohe T. High ambient temperature dampens adaptive immune responses to influenza A virus infection. Proc Natl Acad Sci U S A. 2019 Feb 19;116(8):3118-3125. |
地球温暖化は、さまざまな感染症を媒介する生物(ジカウイルスを媒介する蚊や重症熱性血小板減少症候群ウイルス(SFTSウイルス)を媒介するマダニ等)の生息域を拡大させますが、外気温がウイルス感染後に誘導される免疫応答に与える影響については不明でした。また腸内細菌叢がインフルエンザウイルスに対する免疫応答の誘導に役立つ理由も未解明のままでした。今回、地球温暖化を想定した36℃という暑い環境で飼育したマウスは、22℃で飼育したマウスと比較して、インフルエンザウイルス、ジカウイルス、SFTSウイルスの感染後に誘導される免疫応答が低下することを見出しました。暑い環境で飼育したマウスは摂食量が低下しており、この摂食量の低下が免疫応答の低下につながる要因のひとつでした。そこで、宿主の栄養状態がインフルエンザウイルスに対する免疫応答の誘導に重要な役割を果たしているという仮説を立てて検証したところ、36℃で飼育したマウスに腸内細菌由来代謝産物(酪酸、プロピオン酸、酢酸)やグルコースを投与すると、低下していたウイルス特異的な免疫応答が部分的に回復することを見出しました。以上の成果は、外気温がウイルス特異的な免疫応答の誘導に影響を与えることを示した世界で初めての例であり、腸内細菌叢がインフルエンザウイルス特異的な免疫応答に役立つ理由を解明した極めて重要な知見です。
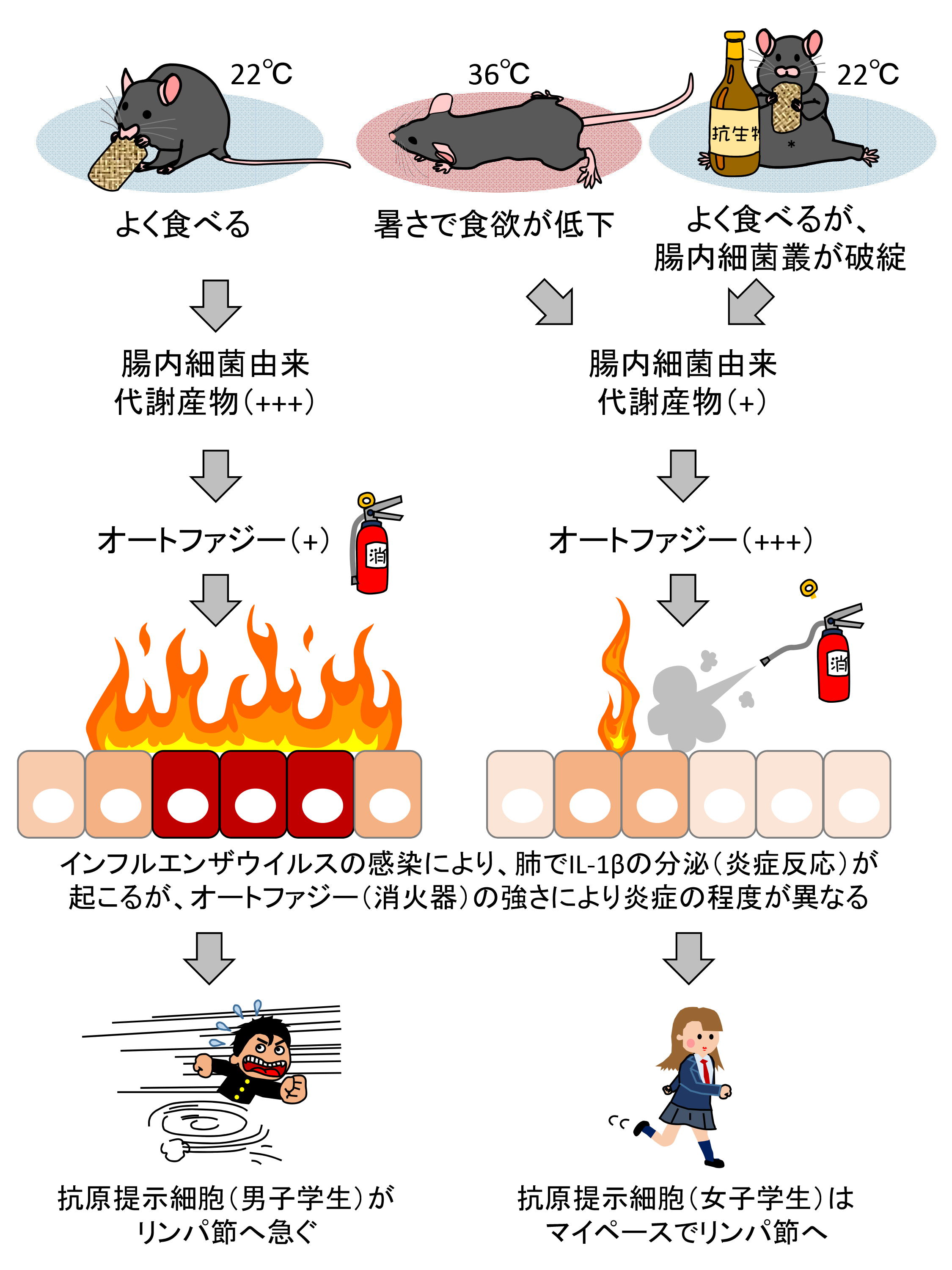
図.本研究成果のまとめ(予測モデル)
通常、よく食べることと健康な腸内細菌の働きにより、腸内細菌由来代謝産物(短鎖脂肪酸など)が多く作られる。一方、暑さによる食欲の低下や、抗生物質による腸内細菌叢のバランスの破綻は、腸内細菌叢由来代謝産物の産生を低下させると考えられる。腸内細菌叢由来代謝産物やグルコースなどは、血流に乗り、肺のオートファジー(図中の消火器)の強さを変化させる。インフルエンザウイルスが感染すると炎症反応に関わるIL-1βが産生されるが、オートファジー(図中の消火器)の強さにより、この炎症(図中の炎)の程度が異なる。IL-1βの産生(炎症)が強いと、肺の抗原提示細胞(図中の男子学生)は、リンパ節へと急ぎ、免疫応答の誘導を助ける。IL-1βの産生(炎症)が低いと、肺の抗原提示細胞(図中の女子学生)は、マイペースでリンパ節へ向かうため、免疫細胞への連絡が遅れる。
・プレスリリース
・UTokyo FOCUS
・東京大学環境報告書2019
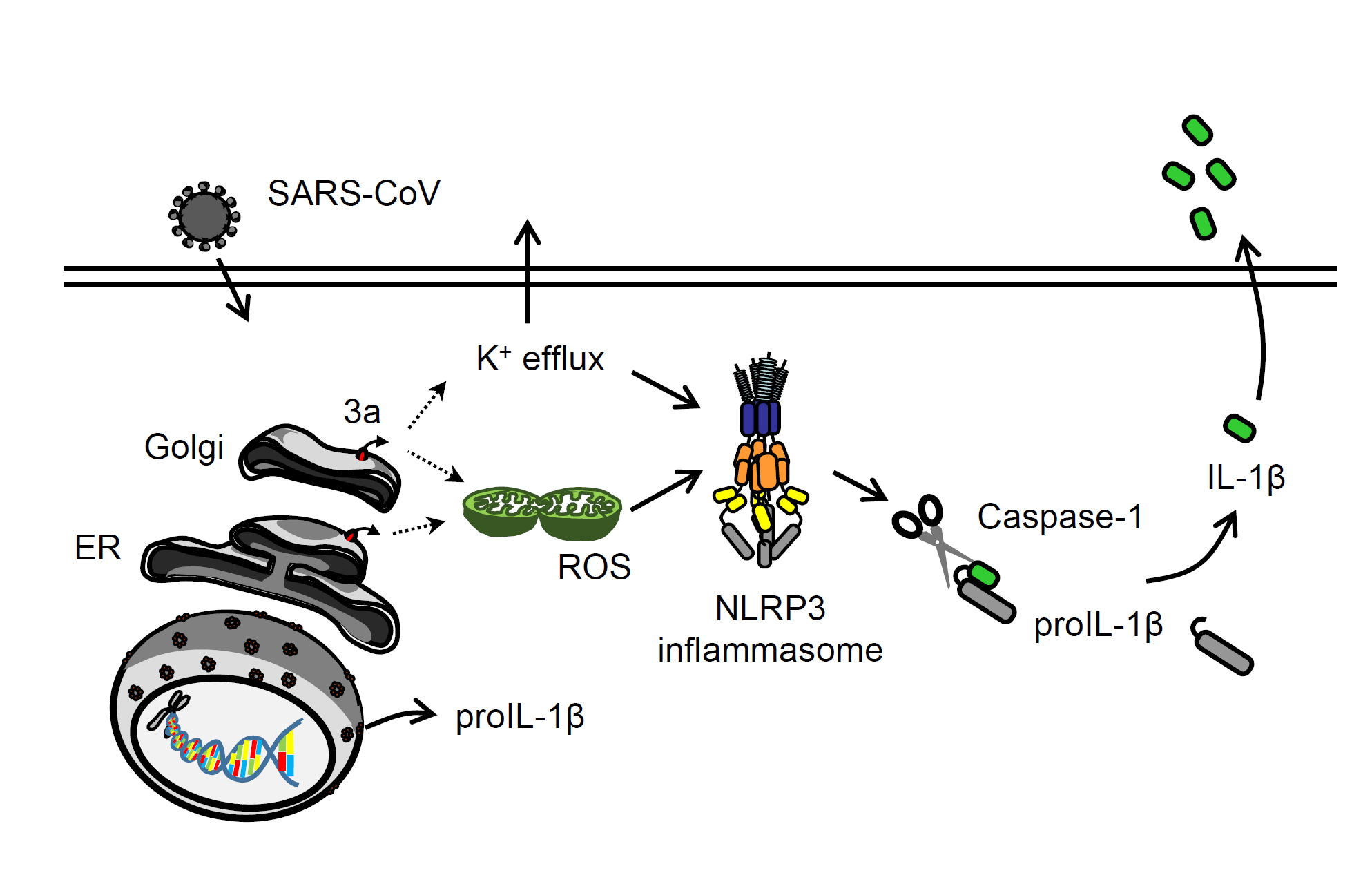
| Chen IY, Moriyama M, Chang MF, Ichinohe T. Severe Acute Respiratory Syndrome Coronavirus Viroporin 3a Activates the NLRP3 Inflammasome. Front Microbiol. 2019 Jan 29;10:50. |
重症急性呼吸器症候群(Severe acute respiratory syndrome; SARS)は、SARSコロナウイルス (SARS-CoV) によって引き起こされる呼吸器感染症です。2002年のアウトブレイクでは香港を中心に8,000人以上が感染し、774人が死亡したとされています(致命率9.6%)。SARS-CoVがヒトの呼吸器に感染すると炎症により重症の肺炎を引き起こします。そのためSARS-CoVが炎症を起こすメカニズムを解明することはウイルスの病原性の理解に重要です。今回、SARS-CoVの3aタンパク質が、この炎症反応(IL-1βの産生)に関わっていることを明らかにしました。具体的には3aタンパク質を細胞に発現させるとそれだけでIL-1βの産生が認められ、3aタンパク質のイオンチャネル活性に必要であるとされる127、130、133番目のシステインをセリンに置換した変異3aタンパク質では、IL-1βの産生能が消失することを見出しました。また3aタンパク質は小胞体(ER)やゴルジ体(Golgi)に局在し、細胞内K+の流出やミトコンドリアの活性酸素種(ROS)の産生を引き起こすことにより、NLRP3 inflammasome依存的なIL-1βの産生(炎症反応)を誘導していることが分かりました。以上の成果は、SARS-CoVが重篤な肺炎を引き起こすというウイルスの病原性の理解や、ウイルス感染による過剰な炎症を抑える治療薬の開発などに役立つと期待されます。
参考:論文引用回数
重症熱性血小板減少症候群(Severe fever with thrombocytopenia syndrome, SFTS)ウイルスは、2011年に中国の研究者らによって発見された新しいウイルスです(Yu et al. N Engl J Med. 2011)。日本でも2013年ごろから西日本を中心に感染者が確認されています。SFTSウイルスの主な感染経路は、森林や草地に生息するマダニを介したものですが、最近では衰弱した野良猫を介抱しようとした女性が、猫に噛まれてSFTSを発症し死亡したというニュースが大きな話題となりました。SFTSに感染した際の致死率は10〜30%とされており、日本における死亡者はすべて50歳代以上で、高齢者が重症化しやすいと考えられています。2013年から2018年6月までに日本国内でも約350人の患者が報告されていますが、これまでのところSFTSウイルスに対する有効な治療薬はなく対症療法しかありません。
マウスを用いた研究では、I型インターフェロン(IFN)受容体欠損マウスが重症のSFTS様症状を呈することから(Liu et al. J Virol. 2014)、SFTSウイルスに対する感染防御には宿主のIFN応答が重要であると考えられています。しかしSFTSウイルスの非構造タンパク質であるNSsタンパク質が、細胞質中に特徴的なの封入体構造を形成してその中に自然免疫関連タンパク質(TBK1など)を隔離することにより(図A)、宿主のIFN応答を抑制することでウイルスの増殖に有利な環境をつくりだしています(Wu et al. J Virol. 2014, Santiago et al. J Virol. 2014)。本研究では、SFTSウイルスのNSsタンパク質が宿主のIFN応答を抑制するのに必要な機能ドメインを同定することを目的に実験をスタートさせました。するとNSsタンパク質中でβシートを形成すると予測された21番目のバリンと23番目のロイシンが、NSsとTBK1との相互作用および宿主のIFN応答の抑制に必須であることが明らかとなりました(図B)。さらにNSsタンパク質は、TBK1のキナーゼドメイン(N末端から307番目までのアミノ酸)と相互作用して、宿主のIFN応答に必要なTBK1のリン酸化を抑制していることを明らかにしました。これまでSFTSウイルスのNSsタンパク質によるIFN応答の抑制には、NSsが形成する細胞内の封入体構造が必要であると考えられていましたが、今回、NSsタンパク質によるこの封入体形成はIFN応答の抑制には必ずしも必要ではなく、TBK1との相互作用そのものが必要であることが明らかとなりました(封入体構造を形成しないNSsの変異体でもTBK1と相互作用することができれば宿主のIFN応答を抑制できた)。本研究成果は、SFTSウイルスに対する新しい治療薬の開発やSFTSウイルスの病原性を理解するのに役立つ極めて重要な知見であると言えます。
.jpg)
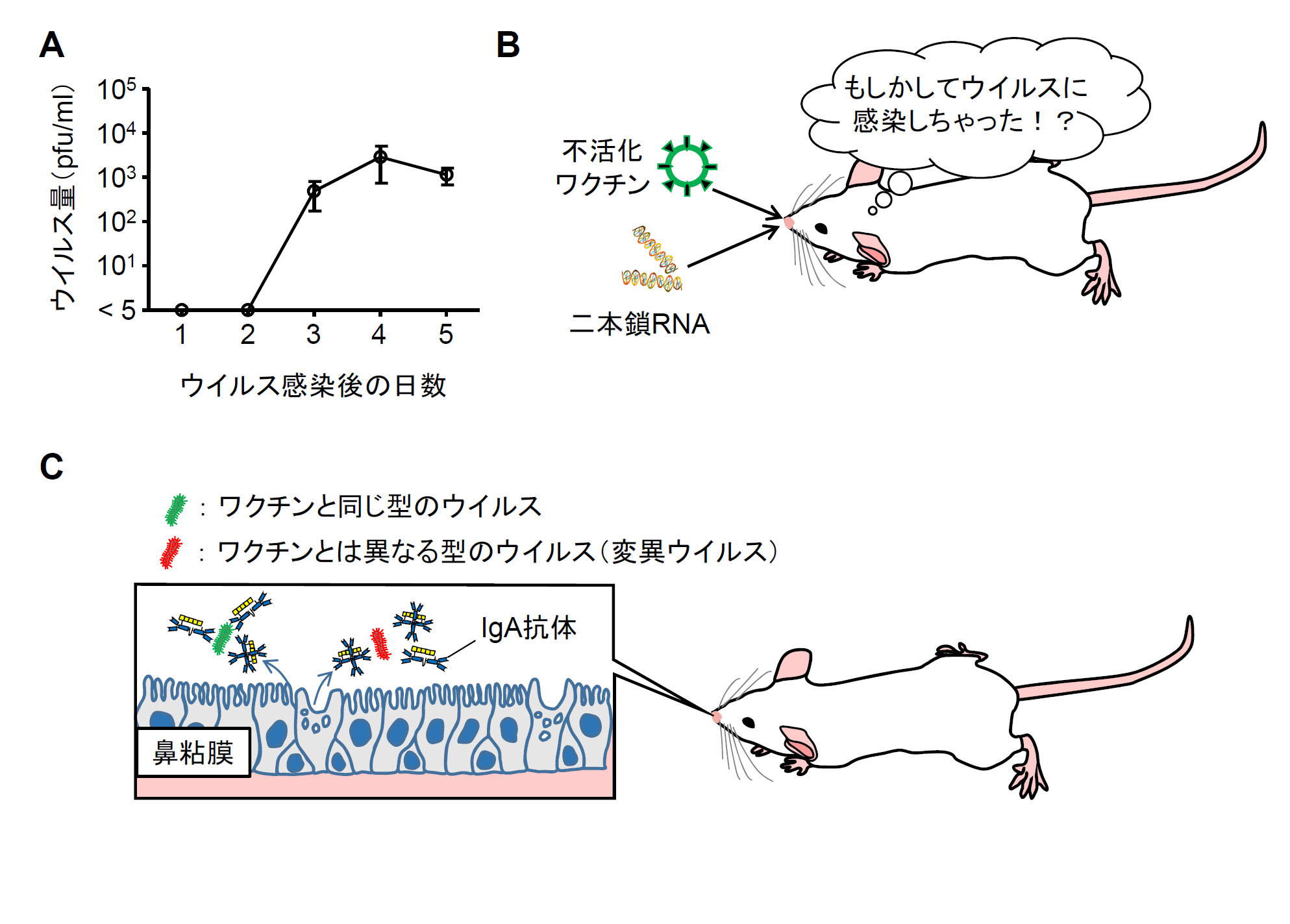
インフルエンザに罹患すると変異ウイルスに対しても有効な鼻粘膜上の粘膜免疫応答(ウイルス特異的なIgA抗体)が誘導されます。従って、効果的なインフルエンザワクチンの開発には、鼻粘膜上におけるインフルエンザウイルスの感染を模倣(マネ)することが必要です。今回、我々はインフルエンザウイルスがマウスの鼻粘膜で増殖する場合、ウイルス量のピークが感染5日目までであることに着目しました(図A)。そこでインフルエンザワクチンとウイルスRNAを模倣した二本鎖RNAを混ぜて、5日間連続でマウスに経鼻投与すると(図B)、実際にインフルエンザウイルスに感染したときと同じような鼻粘膜上のウイルス特異的なIgA抗体が誘導されることが明らかとなりました。このウイルス特異的な鼻粘膜上のIgA抗体は、ワクチン接種から少なくとも半年間持続することも分かりました。ワクチン接種を受けたマウスは、ワクチンと同じ型のウイルスだけでなく、変異ウイルスに対しても感染防御効果を示します(図C)。これらの成果は、私たちが免疫を持っていない新しいウイルス(流行の予測が困難な新型インフルエンザウイルスやワクチンを受けたことがない小児にとっては毎年流行するインフルエンザウイルス)に対して、迅速かつ効率よく防御免疫を誘導するワクチンの開発に役立つ可能性があります。
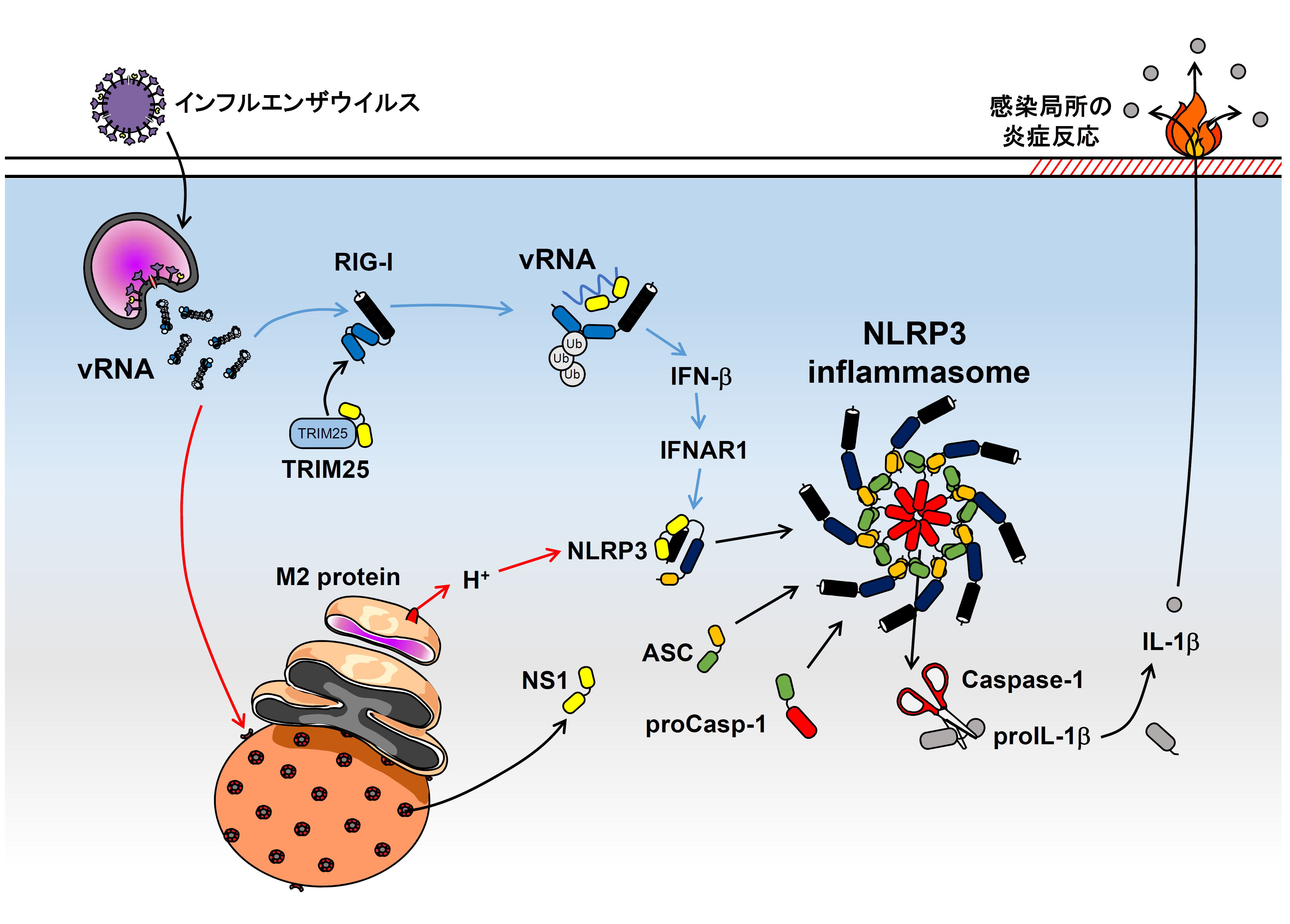
インフルエンザウイルスのM2タンパク質は、トランスゴルジ中のH+を細胞質中へ流出させることにより、細胞質中の自然免疫受容体であるNLRP3 inflammasomeを活性化させています(図の赤矢印、Ichinohe et al. Nat Immunol. 2010)。またRIG-I経路によるインターフェロン応答はNLRP3 inflammasomeの活性化を増大させているという報告もあります(図の青矢印、Pothlichet et al. PLoS Pathog. 2013)。このようなインフルエンザウイルス感染によるinflammasomeの活性化とIL-1βの分泌は、感染局所の炎症反応だけでなく、その後のウイルス特異的な免疫応答に必要です(Ichinohe et al. J Exp Med. 2009, Pang et al. Nat Immunol. 2013)。今回、インフルエンザウイルスのNS1タンパク質がNLRP3と相互作用することにより、NLRP3 inflammasomeの活性化とそれに続くIL-1βの産生を抑制していることを明らかにしました。この抑制効果には、RNA結合ドメイン(38番目のアルギニンと41番目のリシン)と、TRIM25結合ドメイン(96, 97番目のグルタミン酸)が必要であったことから、ウイルスRNAによるNLRP3 inflammasomeの活性化経路(青矢印)にもこのNS1タンパク質が大きく関与していることが明らかとなりました。本研究成果は、インフルエンザウイルスの病原性を理解するのにとても重要です。
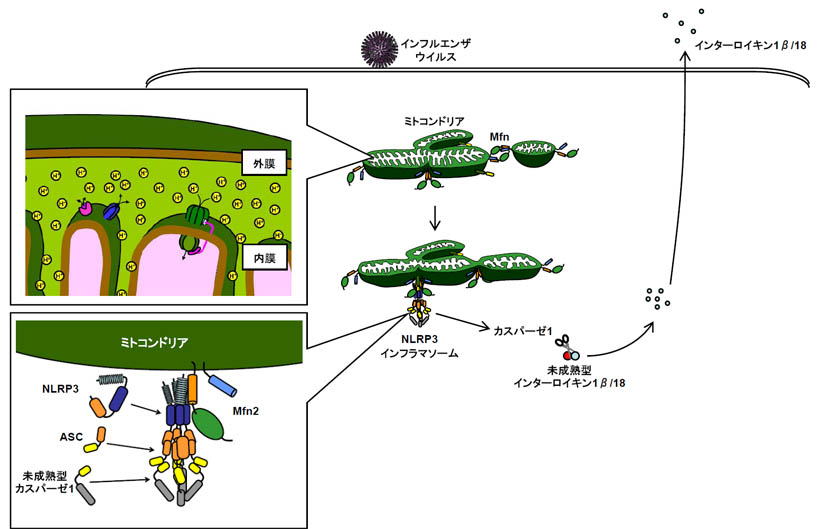
インフルエンザウイルスが細胞に感染すると、細胞内のタンパク質複合体であるNLRP3 inflammasomeが活性化して炎症反応を引き起こします。この炎症反応はインフルエンザウイルス特異的免疫応答の誘導に必要です。今回、インフルエンザウイルスが感染すると細胞内のNLRP3が、ミトコンドリアの外膜タンパク質であるmitofusin 2(Mfn2)に結合することを明らかにしました。このNLRP3とMfn2の結合は、ミトコンドリアの膜電位(ミトコンドリア内膜における局所的な電気的勾配)に依存的で、インフルエンザウイルス感染によるNLRP3 inflammsomeの活性化と炎症誘発性サイトカイン(IL-1β)の産生に必要でした。このことは、ウイルス感染後の過剰な炎症を抑えるような治療薬の開発、または炎症を起こさせることによりインフルエンザワクチンの効果を高めるような物質(アジュバント)の開発に役立つと期待されます。
・プレスリリース
・UTokyo FOCUS
(2013年10月22日。日本経済新聞社が記事利用を承諾しています。許諾番号30037956)