Division of Cellular and Molecular Biology
The Institute of Medical Science
The University of Tokyo
The Institute of Medical Science
The University of Tokyo
TRAF偺榖丂戞嶰夞
IL-1僔僌僫儖偵偍偗傞TRAF6偵傛傞擇憡惈NF-κB妶惈壔儌僨儖偺採彞
Kohsuke Yamazaki, Jin Gohda, Atsuhiro Kanayama, Yusei Miyamoto, Hiroaki Sakurai, Masahiro Yamamoto, Shizuo Akira, Hidetoshi Hayashi, Bing Su, Jun-ichiro Inoue.
Science Signaling 2, Issue 93, ra66, October 2009
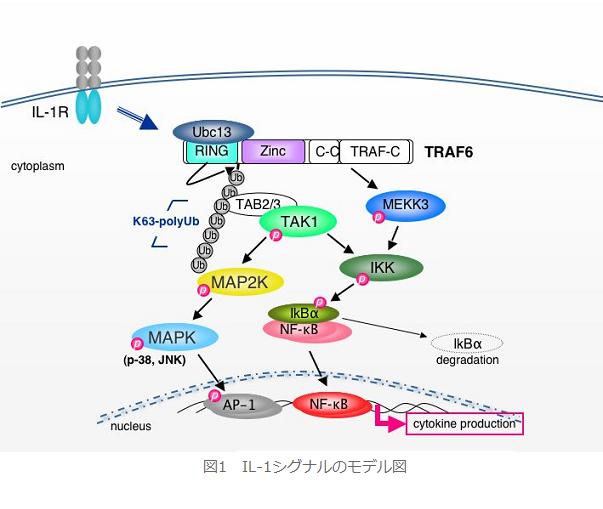
丂嵶朎枌忋偵敪尰偟偰偄傞IL-1庴梕懱偵IL-1偑寢崌偡傞偙偲偵傛傝嵶朎撪偱僔僌僫儖揱払偑桿摫偝傟丆妶惈壔偝傟偨揮幨場巕NF-κB偵傛偭偰丆僒僀僩僇僀儞傗働儌僇僀儞偑敪尰桿摫偝傟傑偡丏偦偺僔僌僫儖揱払乮恾侾乯偵偍偄偰丆儐價僉僠儞儕僈乕僛妶惈乮E3妶惈乯傪桳偡傞RING僪儊僀儞傪帩偮傾僟僾僞乕暘巕TRAF6偑廳梫側栶妱傪偟偰偍傝丆傑偩枹摨掕偱偡偑儐價僉僠儞寢崌峺慺乮E2乯傪夘偟偰帺屓傪K63宆億儕儐價僉僠儞壔偟傑偡丏偙偺K63宆億儕儐價僉僠儞嵔偼僾儘僥傾僜乕儉暘夝偺昗揑偱偼側偔丆僔僌僫儖暘巕偑暋崌懱宍惉偡傞偨傔偺懌応偵側傞偲峫偊傜傟偰偍傝丆壓棳偺儕儞巁壔峺慺偱偁傞TAK1偲TAB2偺暋崌懱偑偙偺億儕儐價僉僠儞嵔傪夘偟偰TRAF6偲寢崌偡傞偙偲偑抦傜傟偰偄傑偡丏偙偺寢崌偵傛傝妶惈壔偟偨TAK1偑IKK 傪儕儞巁壔偡傞偙偲偱妶惈壔偝偣丆懕偄偰IKK偑NF-κB偺梷惂場巕偱偁傞IκBα偺儕儞巁壔丆偦傟偵懕偔僾儘僥傾僜乕儉暘夝傪桿摫偡傞偙偲偱丆NF-κB偺妀堏峴傪懀偟丆揮幨妶惈傪桿摫偡傞偲峫偊傜傟偰偄傑偡丏堦曽丆TRAF6偺壓棳偵埵抲偡傞儕儞巁壔峺慺偱偁傞MEKK3傕TAK1偲摨條偵IL-1僔僌僫儖偵娭梌偟偰偍傝丆IKK傪儕儞巁壔偡傞偙偲偱NF-κB妶惈傪懀偡偙偲偑抦傜傟偰偄傑偡丏偝傜偵丆偙偺TAK1偲MEKK3偼嫟偵MAPK妶惈偵傕昁恵偱偁傞偙偲偑夝偭偰偍傝丆懕偄偰AP-1偺妶惈壔傪桿摫偟傑偡丏偟偐偟側偑傜丆TAK1偲MEKK3丆偦偟偰TRAF6偺3幰偼IL-1僔僌僫儖偵怺偔娭梌偡傞偵傕娭傢傜偢丆TRAF6壓棳偱偺TAK1偲MEKK3偺暔棟揑憡屳嶌梡傗婡擻揑娭學偵偮偄偰偼柧傜偐偵側偭偰偄傑偣傫偱偟偨丏杮尋媶偱偼丆2偮偺儕儞巁壔峺慺乮TAK1偲MEKK3乯偺妶惈壔婡峔偺夝愅傪拞怱偲偟偰恑傔偨寢壥丆TRAF6偵傛傞岻柇側NF-κB妶惈壔偺惂屼婡峔偑柧傜偐偵側傝傑偟偨丏
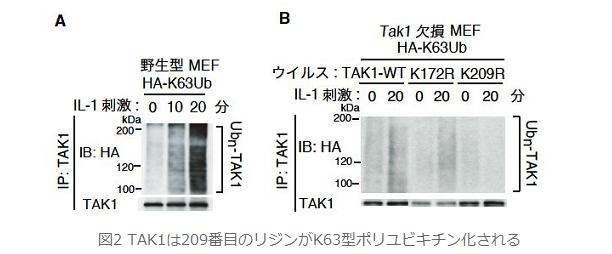
丂TAK1偺億儕儐價僉僠儞壔偺婡擻傪柧傜偐偵偡傞偨傔丆TAK1偺摿堎揑側億儕儐價僉僠儞壔晹埵偺摨掕傪帋傒傑偟偨丏懡偔偺庬娫偱曐懚偝傟偰偄傞7偮偺儕僕儞傪拪弌偟丆偦傟偧傟偺儕僕儞傪傾儖僊僯儞偵揰曄堎偝偣偨7庬椶偺TAK1 K/R曄堎懱傪峔抸偟傑偟偨丏偦傟傜傪栰惗宆偺MEF偲摨掱搙偺TAK1敪尰検偵側傞條偵挷惍偟偰丆TAK1偺寚懝MEF偵埨掕敪尰偝偣丆偝傜偵HA-K63Ub傪埨掕敪尰偝偣傞偙偲偱丆惗棟揑忦審壓偵偍偗傞IL-1僔僌僫儖埶懚揑側TAK1偺K63宆偺億儕儐價僉僠儞壔傪専弌偟傑偟偨丏偡傞偲丆209斣栚偺儕僕儞偵揰曄堎傪梌偊偨TAK1乮TAK1 K209R乯偺傒偵偍偄偰丆TAK1偺億儕儐價僉僠儞壔偑挊偟偔尭庛偟傑偟偨乮恾俀俛乯丏偙傟傛傝丆IL-1僔僌僫儖埶懚揑側TAK1偺K63宆億儕儐價僉僠儞壔偼K209偑廋忺晹埵偱偁傞偙偲偑暘偐傝傑偟偨丏
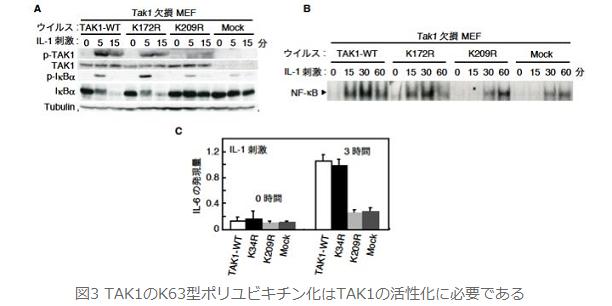
丂TAK1偺億儕儐價僉僠儞壔偺NF-κB妶惈壔傊偺娭梌傪柧傜偐偵偡傞偨傔丆TAK1寚懝MEF偵TAK1偺栰惗宆傪敪尰偝偣偨嵶朎乮TAK1 WT MEF乯偲儐價僉僠儞壔偝傟側偄曄堎懱 TAK1 K209R傪TAK1寚懝MEF偵敪尰偝偣偨嵶朎乮TAK1 K209R MEF乯偵懳偟偰丆IL-1巋寖傪梌偊傑偟偨丏TAK1 WT NEF偲斾妑偟偰TAK1 K209R MEF偱偼丆TAK1偺妶惈壔乮恾俁俙乯丆NF-κB偺DNA寢崌擻椡乮恾俁俛乯丆偝傜偵偼丆NF-κB妶惈偺昗揑堚揱巕偱偁傞IL-6偺mRNA敪尰検乮恾俁俠乯傕挊偟偔尭庛偟傑偟偨丏偙傟傜偺寢壥偼丆Ubc13乮E2乯偲TRAF6乮E3乯傪夘偟偰惗惉偝傟傞TAK1偺億儕儐價僉僠儞壔偼TAK1帺恎偺妶惈壔偲偦傟偵懕偔NF-κB偺廫暘側妶惈傗IL-6嶻惗偵昁梫偱偁傞偙偲偑帵偝傟傑偟偨丏
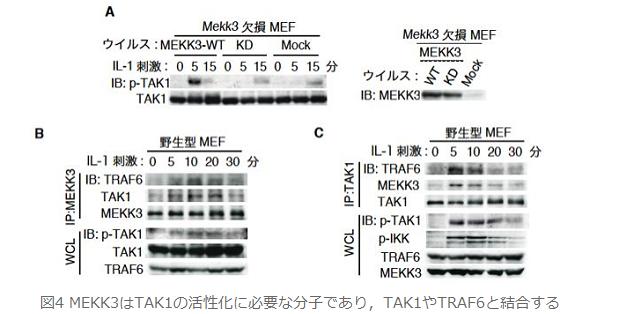
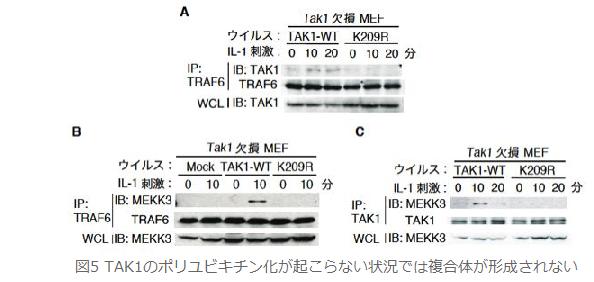
丂偝偰丆偙偺3暘巕偺暋崌懱宍惉偵TAK1偺億儕儐價僉僠儞壔偑偳偺傛偆偵娭梌偡傞偐傪丆TAK1 WT MEF偲TAK1 K209R MEF傪梡偄偰専摙偟傑偟偨丏嬃偔傋偒偙偲偵丆TAK1 WT MEF偱妋擣偝傟偨TRAF6偲TAK1偺寢崌乮恾俆俙乯丆TRAF6偲MEKK3偺寢崌 乮恾俆俛乯丆偦偟偰TAK1偲MEKK3偺寢崌乮恾俆俠乯偑TAK1 K209R MEF偱偼杦偳徚幐偟傑偟偨丏偙傟傜偺寢壥偐傜TAK1偺億儕儐價僉僠儞壔偼TRAF6丆TAK1丆MEKK3偐傜峔惉偝傟傞IL-1僔僌僫儖埶懚揑側暋崌懱宍惉偵昁梫偱偁傞偙偲偑帵偝傟傑偟偨丏偦偟偰丆偙偺暋崌懱宍惉偵傛傝丆MEKK3偵傛傞TAK1偺妶惈壔偑懀偝傟丆懕偄偰NF-κB妶惈壔偑摫偐傟傞偙偲偑帵嵈偝傟傑偟偨丏
丂杮尋媶偵偍偄偰偼丆偙偺Zinc宱楬偺暘巕婡峔偲惗棟婡擻偵偮偄偰偺夝愅傪帋傒傑偟偨丏傑偢丆Zinc宱楬偵娭梌偡傞暘巕傪摨掕偡傞偨傔丆TRAF6 儮R MEF偵懳偟偰siRNA傪梡偄偨TAK1丆枖偼MEKK3偺堚揱巕敪尰梷惂傪偟偨忋偱丆IL-1僔僌僫儖埶懚揑側NF-κB妶惈傪専摙偟偨偲偙傠丆MEKK3偺敪尰梷惂偵傛傝巆懚偟偰偄偨NF-kB妶惈偼姰慡偵徚幐偟傑偟偨偑丆TAK1偺敪尰梷惂偱偼傎偲傫偳曄壔偑偁傝傑偣傫偱偟偨丏偙傟傛傝Zinc宱楬偵偼TAK1偱偼側偔丆MEKK3偑娭梌偟偰偄傞偙偲偑柧傜偐偲側傝傑偟偨丏
丂師偵偦偺惗棟婡擻傪柧傜偐偵偡傞偨傔丆Zinc宱楬偺傒偑婡擻偟側偄TRAF6曄堎懱偺峔抸傪帋傒傑偟偨丏TRAF6偼5偮偺Zinc 僪儊僀儞傪帩偮偲峫偊傜傟偰偄傑偡偑丆N枛抂懁偐傜悢偊偰5斣栚偺Zinc僪儊僀儞偺婡擻傪捵偟偨曄堎懱乮TRAF6 mZ5乯傪TRAF6寚懝MEF偵埨掕敪尰偝偣偨嵶朎乮TRAF6 mZ5 MEF乯偱偼丆TAK1偺妶惈壔偵塭嬁傪梌偊側偄偑丆NF-κB妶惈偺尭庛偑擣傔傜傟丆偝傜偵TAK1偺堚揱巕敪尰梷惂偵傛傝丆NF-κB妶惈偼姰慡偵徚幐偟傑偟偨丏廬偭偰丆TRAF6 mZ5偼RING宱楬偵娭梌偡傞TAK1偺妶惈偵偼曄壔傪梌偊側偄偑丆Zinc宱楬偼婡擻偟側偄曄堎懱偱偁傞偙偲傪帵偟偰偄傑偡丏
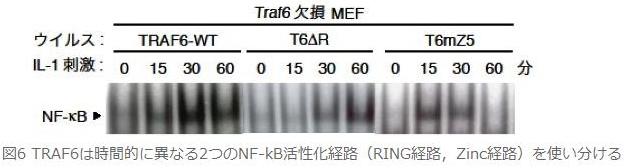
丂偦偟偰丆栰惗宆偺TRAF6偲忋弎偟偨2庬椶偺TRAF6曄堎懱傪TRAF6寚懝MEF偵埨掕敪尰偝偣偨嵶朎乮TRAF6 WT MEF丆TRAF6 儮R MEF丆TRAF6 mZ5 MEF乯偵偍偗傞EMSA傪梡偄偨NF-κB偺DNA寢崌擻椡傪斾妑偟傑偟偨丏TRAF6 WT MEF偵偍偄偰偼丆NF-κB偺DNA寢崌擻椡偼IL-1巋寖屻15暘偱妋擣偱偒丆30暘偱僺乕僋傪寎偊丆60暘偱傗傗尭庛偟傑偡偑丆TRAF6 儮R MEF偱偼丆30暘屻偱弶傔偰妋擣偝傟丆60暘屻偵憹戝偟傑偟偨丏堦曽丆TRAF6 mZ5 MEF偱偼丆15暘屻偱妋擣偝傟丆30暘屻偱傕憹戝偣偢丆60暘屻偵偼徚幐偟偰偄傑偟偨乮恾俇乯丏TRAF6 儮R MEF偲TRAF6 mZ5 MEF偵偍偗傞NF-κB偺DNA寢崌擻椡傪壛嶼偝偣偨傕偺偑丆偪傚偆偳TRAF6 WT MEF偺傕偺偲堦抳偟偰偄傑偟偨丏偙傟傛傝丆RING宱楬偼帪娫揑偵憗偄NF-κB妶惈傪扴摉偟丆堦曽Zinc宱楬偼帪娫揑偵抶偄NF-κB妶惈傪扴摉偡傞偙偲偑帵偝傟丆椉曽偺宱楬偼婡峔揑乮儐價僉僠儞壔傪敽偆偐斲偐乯偵堎側傞偩偗偱偼側偔丆帪娫揑乮RING宱楬偑憗偔丆Zinc宱楬偑抶偄乯偵傕堎側偭偨NF-κB妶惈宱楬偱偁傞偙偲偑帵偝傟傑偟偨丏
丂
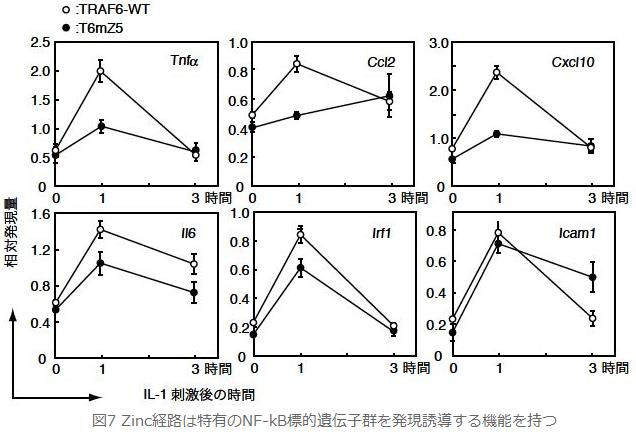
丂偝傜偵丆Zinc宱楬偺惗棟婡擻傪棟夝偡傞偨傔偵NF-κB昗揑堚揱巕偺敪尰偵拝栚偟傑偟偨丏Real-time PCR傪梡偄偰IL-1巋寖屻偺NF-κB昗揑堚揱巕偺mRNA検傪斾妑専摙偟偨偲偙傠丆TRAF6-WT MEF偲斾傋偰TRAF6-mZ5 MEF偱偼IL-6丆IRF1偺敪尰検偼嬐偐側尭彮偵棷傑傝傑偟偨偑丆TNFa丆CCL2丆CXCL10偺敪尰検偼尠挊側尭彮偑尒傜傟傑偟偨丏偙傟傜偺寢壥偐傜丆Zinc宱楬偑堦孮偺NF-κB昗揑堚揱巕偺敪尰桿摫偵廳梫側栶妱傪壥偨偟偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丏NF-κB昗揑堚揱巕偺拞偵偼丆偦偺敪尰偑MAPK偺1偮偱偁傞p38偵埶懚揑側NF-kB昗揑堚揱巕孮乮IL-6丆IRF1丆ICAM1乯偲埶懚惈偺掅偄堚揱巕孮乮TNFa丆CCL2乯偑懚嵼偡傞偙偲偑嵟嬤曬崘偝傟傑偟偨丏傑偨杮尋媶偵偍偄偰丆TRAF6-mZ5 MEF偱偼NF-κB妶惈偵尭庛偑尒傜傟傑偡偑丆p38偺妶惈偵偼杦偳曄壔傪梌偊傑偣傫偱偟偨丏偳偆傗傜丆Zinc宱楬偼p38偵埶懚惈偺掅偄 NF-κB昗揑堚揱巕偺敪尰桿摫偵偍偄偰婡擻偟偰偄傞傛偆偵巚傢傟傑偡偑丆寢榑偡傞偵偼峏側傞夝愅偑昁梫偱偡丏
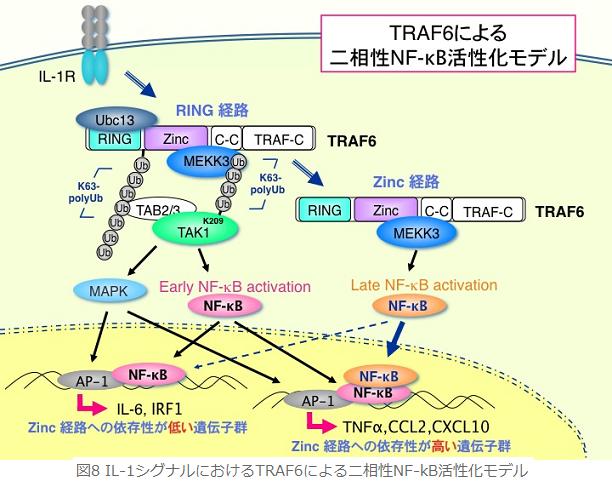
丂崱屻偺壽戣偲偟偰偼丆TAK1偺億儕儐價僉僠儞壔偑暋崌懱宍惉傪桿摫偡傞暘巕婡峔偺夝柧偑嫇偘傜傟傑偡丏億儕儐價僉僠儞壔偼僔僌僫儖暘巕偺暋崌懱宍惉偺偨傔偺懌応偲側傞偲峫偊傜傟偰偄傞偨傔丆TAK1偺億儕儐價僉僠儞嵔偵MEKK3傗TRAF6摍偺僔僌僫儖暘巕偑寢崌偡傞偙偲偵傛傝丆暋崌懱偑埨掕壔偟丆僔僌僫儖偑憹嫮偝傟傞婡峔傪峫偊偰偄傑偡丏傑偨丆RING宱楬偐傜Zinc宱楬傊偺僗僀僢僠儞僌丒僔僗僥儉偺夝柧傕廳梫壽戣偺侾偮偱偡丏
丂IL-1僔僌僫儖偼墛徢斀墳傗姶愼杊屼側偳偺柶塽斀墳丆偝傜偵偼娻偺埆惈壔偵傕婑梌偡傞偙偲偑抦傜傟偰偍傝丆堛椕傊偺墳梡偺柺偱傕戝帠側僔僌僫儖偱偡丏偙偙偱婰偟偨傛偆偵IL-1僔僌僫儖偵偍偄偰TRAF6偼尞偲側傞暘巕偱偁傞偨傔丆TRAF6偵傛傞僔僌僫儖揱払婡峔傪偝傜偵徻嵶偵夝柧偡傞偙偲偱丆偦偺抦尒傪婎斦偲偟偨堛椕傊偺墳梡偑婜懸偝傟傑偡丏
Kohsuke Yamazaki, Jin Gohda, Atsuhiro Kanayama, Yusei Miyamoto, Hiroaki Sakurai, Masahiro Yamamoto, Shizuo Akira, Hidetoshi Hayashi, Bing Su, Jun-ichiro Inoue.
Science Signaling 2, Issue 93, ra66, October 2009
- IL-1僔僌僫儖偼墛徢斀墳傗柶塽墳摎傪桿摫偡傞
丂嵶朎枌忋偵敪尰偟偰偄傞IL-1庴梕懱偵IL-1偑寢崌偡傞偙偲偵傛傝嵶朎撪偱僔僌僫儖揱払偑桿摫偝傟丆妶惈壔偝傟偨揮幨場巕NF-κB偵傛偭偰丆僒僀僩僇僀儞傗働儌僇僀儞偑敪尰桿摫偝傟傑偡丏偦偺僔僌僫儖揱払乮恾侾乯偵偍偄偰丆儐價僉僠儞儕僈乕僛妶惈乮E3妶惈乯傪桳偡傞RING僪儊僀儞傪帩偮傾僟僾僞乕暘巕TRAF6偑廳梫側栶妱傪偟偰偍傝丆傑偩枹摨掕偱偡偑儐價僉僠儞寢崌峺慺乮E2乯傪夘偟偰帺屓傪K63宆億儕儐價僉僠儞壔偟傑偡丏偙偺K63宆億儕儐價僉僠儞嵔偼僾儘僥傾僜乕儉暘夝偺昗揑偱偼側偔丆僔僌僫儖暘巕偑暋崌懱宍惉偡傞偨傔偺懌応偵側傞偲峫偊傜傟偰偍傝丆壓棳偺儕儞巁壔峺慺偱偁傞TAK1偲TAB2偺暋崌懱偑偙偺億儕儐價僉僠儞嵔傪夘偟偰TRAF6偲寢崌偡傞偙偲偑抦傜傟偰偄傑偡丏偙偺寢崌偵傛傝妶惈壔偟偨TAK1偑IKK 傪儕儞巁壔偡傞偙偲偱妶惈壔偝偣丆懕偄偰IKK偑NF-κB偺梷惂場巕偱偁傞IκBα偺儕儞巁壔丆偦傟偵懕偔僾儘僥傾僜乕儉暘夝傪桿摫偡傞偙偲偱丆NF-κB偺妀堏峴傪懀偟丆揮幨妶惈傪桿摫偡傞偲峫偊傜傟偰偄傑偡丏堦曽丆TRAF6偺壓棳偵埵抲偡傞儕儞巁壔峺慺偱偁傞MEKK3傕TAK1偲摨條偵IL-1僔僌僫儖偵娭梌偟偰偍傝丆IKK傪儕儞巁壔偡傞偙偲偱NF-κB妶惈傪懀偡偙偲偑抦傜傟偰偄傑偡丏偝傜偵丆偙偺TAK1偲MEKK3偼嫟偵MAPK妶惈偵傕昁恵偱偁傞偙偲偑夝偭偰偍傝丆懕偄偰AP-1偺妶惈壔傪桿摫偟傑偡丏偟偐偟側偑傜丆TAK1偲MEKK3丆偦偟偰TRAF6偺3幰偼IL-1僔僌僫儖偵怺偔娭梌偡傞偵傕娭傢傜偢丆TRAF6壓棳偱偺TAK1偲MEKK3偺暔棟揑憡屳嶌梡傗婡擻揑娭學偵偮偄偰偼柧傜偐偵側偭偰偄傑偣傫偱偟偨丏杮尋媶偱偼丆2偮偺儕儞巁壔峺慺乮TAK1偲MEKK3乯偺妶惈壔婡峔偺夝愅傪拞怱偲偟偰恑傔偨寢壥丆TRAF6偵傛傞岻柇側NF-κB妶惈壔偺惂屼婡峔偑柧傜偐偵側傝傑偟偨丏
- TAK1偺K63宆億儕儐價僉僠儞壔偼TAK1偺妶惈壔偵昁梫偱偁傞
丂TAK1偺億儕儐價僉僠儞壔偺婡擻傪柧傜偐偵偡傞偨傔丆TAK1偺摿堎揑側億儕儐價僉僠儞壔晹埵偺摨掕傪帋傒傑偟偨丏懡偔偺庬娫偱曐懚偝傟偰偄傞7偮偺儕僕儞傪拪弌偟丆偦傟偧傟偺儕僕儞傪傾儖僊僯儞偵揰曄堎偝偣偨7庬椶偺TAK1 K/R曄堎懱傪峔抸偟傑偟偨丏偦傟傜傪栰惗宆偺MEF偲摨掱搙偺TAK1敪尰検偵側傞條偵挷惍偟偰丆TAK1偺寚懝MEF偵埨掕敪尰偝偣丆偝傜偵HA-K63Ub傪埨掕敪尰偝偣傞偙偲偱丆惗棟揑忦審壓偵偍偗傞IL-1僔僌僫儖埶懚揑側TAK1偺K63宆偺億儕儐價僉僠儞壔傪専弌偟傑偟偨丏偡傞偲丆209斣栚偺儕僕儞偵揰曄堎傪梌偊偨TAK1乮TAK1 K209R乯偺傒偵偍偄偰丆TAK1偺億儕儐價僉僠儞壔偑挊偟偔尭庛偟傑偟偨乮恾俀俛乯丏偙傟傛傝丆IL-1僔僌僫儖埶懚揑側TAK1偺K63宆億儕儐價僉僠儞壔偼K209偑廋忺晹埵偱偁傞偙偲偑暘偐傝傑偟偨丏
丂TAK1偺億儕儐價僉僠儞壔偺NF-κB妶惈壔傊偺娭梌傪柧傜偐偵偡傞偨傔丆TAK1寚懝MEF偵TAK1偺栰惗宆傪敪尰偝偣偨嵶朎乮TAK1 WT MEF乯偲儐價僉僠儞壔偝傟側偄曄堎懱 TAK1 K209R傪TAK1寚懝MEF偵敪尰偝偣偨嵶朎乮TAK1 K209R MEF乯偵懳偟偰丆IL-1巋寖傪梌偊傑偟偨丏TAK1 WT NEF偲斾妑偟偰TAK1 K209R MEF偱偼丆TAK1偺妶惈壔乮恾俁俙乯丆NF-κB偺DNA寢崌擻椡乮恾俁俛乯丆偝傜偵偼丆NF-κB妶惈偺昗揑堚揱巕偱偁傞IL-6偺mRNA敪尰検乮恾俁俠乯傕挊偟偔尭庛偟傑偟偨丏偙傟傜偺寢壥偼丆Ubc13乮E2乯偲TRAF6乮E3乯傪夘偟偰惗惉偝傟傞TAK1偺億儕儐價僉僠儞壔偼TAK1帺恎偺妶惈壔偲偦傟偵懕偔NF-κB偺廫暘側妶惈傗IL-6嶻惗偵昁梫偱偁傞偙偲偑帵偝傟傑偟偨丏
- TAK1偺億儕儐價僉僠儞壔偼帺恎偺妶惈壔傪懀偡暋崌懱宍惉傪桿摫偡傞
丂偝偰丆偙偺3暘巕偺暋崌懱宍惉偵TAK1偺億儕儐價僉僠儞壔偑偳偺傛偆偵娭梌偡傞偐傪丆TAK1 WT MEF偲TAK1 K209R MEF傪梡偄偰専摙偟傑偟偨丏嬃偔傋偒偙偲偵丆TAK1 WT MEF偱妋擣偝傟偨TRAF6偲TAK1偺寢崌乮恾俆俙乯丆TRAF6偲MEKK3偺寢崌 乮恾俆俛乯丆偦偟偰TAK1偲MEKK3偺寢崌乮恾俆俠乯偑TAK1 K209R MEF偱偼杦偳徚幐偟傑偟偨丏偙傟傜偺寢壥偐傜TAK1偺億儕儐價僉僠儞壔偼TRAF6丆TAK1丆MEKK3偐傜峔惉偝傟傞IL-1僔僌僫儖埶懚揑側暋崌懱宍惉偵昁梫偱偁傞偙偲偑帵偝傟傑偟偨丏偦偟偰丆偙偺暋崌懱宍惉偵傛傝丆MEKK3偵傛傞TAK1偺妶惈壔偑懀偝傟丆懕偄偰NF-κB妶惈壔偑摫偐傟傞偙偲偑帵嵈偝傟傑偟偨丏
- TRAF6偼婡峔揑丒帪娫揑偵堎側傞俀偮偺宱楬傪梡偄丆NF-κB妶惈傪惛枾偵惂屼偡傞
丂杮尋媶偵偍偄偰偼丆偙偺Zinc宱楬偺暘巕婡峔偲惗棟婡擻偵偮偄偰偺夝愅傪帋傒傑偟偨丏傑偢丆Zinc宱楬偵娭梌偡傞暘巕傪摨掕偡傞偨傔丆TRAF6 儮R MEF偵懳偟偰siRNA傪梡偄偨TAK1丆枖偼MEKK3偺堚揱巕敪尰梷惂傪偟偨忋偱丆IL-1僔僌僫儖埶懚揑側NF-κB妶惈傪専摙偟偨偲偙傠丆MEKK3偺敪尰梷惂偵傛傝巆懚偟偰偄偨NF-kB妶惈偼姰慡偵徚幐偟傑偟偨偑丆TAK1偺敪尰梷惂偱偼傎偲傫偳曄壔偑偁傝傑偣傫偱偟偨丏偙傟傛傝Zinc宱楬偵偼TAK1偱偼側偔丆MEKK3偑娭梌偟偰偄傞偙偲偑柧傜偐偲側傝傑偟偨丏
丂師偵偦偺惗棟婡擻傪柧傜偐偵偡傞偨傔丆Zinc宱楬偺傒偑婡擻偟側偄TRAF6曄堎懱偺峔抸傪帋傒傑偟偨丏TRAF6偼5偮偺Zinc 僪儊僀儞傪帩偮偲峫偊傜傟偰偄傑偡偑丆N枛抂懁偐傜悢偊偰5斣栚偺Zinc僪儊僀儞偺婡擻傪捵偟偨曄堎懱乮TRAF6 mZ5乯傪TRAF6寚懝MEF偵埨掕敪尰偝偣偨嵶朎乮TRAF6 mZ5 MEF乯偱偼丆TAK1偺妶惈壔偵塭嬁傪梌偊側偄偑丆NF-κB妶惈偺尭庛偑擣傔傜傟丆偝傜偵TAK1偺堚揱巕敪尰梷惂偵傛傝丆NF-κB妶惈偼姰慡偵徚幐偟傑偟偨丏廬偭偰丆TRAF6 mZ5偼RING宱楬偵娭梌偡傞TAK1偺妶惈偵偼曄壔傪梌偊側偄偑丆Zinc宱楬偼婡擻偟側偄曄堎懱偱偁傞偙偲傪帵偟偰偄傑偡丏
丂偦偟偰丆栰惗宆偺TRAF6偲忋弎偟偨2庬椶偺TRAF6曄堎懱傪TRAF6寚懝MEF偵埨掕敪尰偝偣偨嵶朎乮TRAF6 WT MEF丆TRAF6 儮R MEF丆TRAF6 mZ5 MEF乯偵偍偗傞EMSA傪梡偄偨NF-κB偺DNA寢崌擻椡傪斾妑偟傑偟偨丏TRAF6 WT MEF偵偍偄偰偼丆NF-κB偺DNA寢崌擻椡偼IL-1巋寖屻15暘偱妋擣偱偒丆30暘偱僺乕僋傪寎偊丆60暘偱傗傗尭庛偟傑偡偑丆TRAF6 儮R MEF偱偼丆30暘屻偱弶傔偰妋擣偝傟丆60暘屻偵憹戝偟傑偟偨丏堦曽丆TRAF6 mZ5 MEF偱偼丆15暘屻偱妋擣偝傟丆30暘屻偱傕憹戝偣偢丆60暘屻偵偼徚幐偟偰偄傑偟偨乮恾俇乯丏TRAF6 儮R MEF偲TRAF6 mZ5 MEF偵偍偗傞NF-κB偺DNA寢崌擻椡傪壛嶼偝偣偨傕偺偑丆偪傚偆偳TRAF6 WT MEF偺傕偺偲堦抳偟偰偄傑偟偨丏偙傟傛傝丆RING宱楬偼帪娫揑偵憗偄NF-κB妶惈傪扴摉偟丆堦曽Zinc宱楬偼帪娫揑偵抶偄NF-κB妶惈傪扴摉偡傞偙偲偑帵偝傟丆椉曽偺宱楬偼婡峔揑乮儐價僉僠儞壔傪敽偆偐斲偐乯偵堎側傞偩偗偱偼側偔丆帪娫揑乮RING宱楬偑憗偔丆Zinc宱楬偑抶偄乯偵傕堎側偭偨NF-κB妶惈宱楬偱偁傞偙偲偑帵偝傟傑偟偨丏
丂
丂偝傜偵丆Zinc宱楬偺惗棟婡擻傪棟夝偡傞偨傔偵NF-κB昗揑堚揱巕偺敪尰偵拝栚偟傑偟偨丏Real-time PCR傪梡偄偰IL-1巋寖屻偺NF-κB昗揑堚揱巕偺mRNA検傪斾妑専摙偟偨偲偙傠丆TRAF6-WT MEF偲斾傋偰TRAF6-mZ5 MEF偱偼IL-6丆IRF1偺敪尰検偼嬐偐側尭彮偵棷傑傝傑偟偨偑丆TNFa丆CCL2丆CXCL10偺敪尰検偼尠挊側尭彮偑尒傜傟傑偟偨丏偙傟傜偺寢壥偐傜丆Zinc宱楬偑堦孮偺NF-κB昗揑堚揱巕偺敪尰桿摫偵廳梫側栶妱傪壥偨偟偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丏NF-κB昗揑堚揱巕偺拞偵偼丆偦偺敪尰偑MAPK偺1偮偱偁傞p38偵埶懚揑側NF-kB昗揑堚揱巕孮乮IL-6丆IRF1丆ICAM1乯偲埶懚惈偺掅偄堚揱巕孮乮TNFa丆CCL2乯偑懚嵼偡傞偙偲偑嵟嬤曬崘偝傟傑偟偨丏傑偨杮尋媶偵偍偄偰丆TRAF6-mZ5 MEF偱偼NF-κB妶惈偵尭庛偑尒傜傟傑偡偑丆p38偺妶惈偵偼杦偳曄壔傪梌偊傑偣傫偱偟偨丏偳偆傗傜丆Zinc宱楬偼p38偵埶懚惈偺掅偄 NF-κB昗揑堚揱巕偺敪尰桿摫偵偍偄偰婡擻偟偰偄傞傛偆偵巚傢傟傑偡偑丆寢榑偡傞偵偼峏側傞夝愅偑昁梫偱偡丏
- 傑偲傔偲崱屻偺壽戣
丂崱屻偺壽戣偲偟偰偼丆TAK1偺億儕儐價僉僠儞壔偑暋崌懱宍惉傪桿摫偡傞暘巕婡峔偺夝柧偑嫇偘傜傟傑偡丏億儕儐價僉僠儞壔偼僔僌僫儖暘巕偺暋崌懱宍惉偺偨傔偺懌応偲側傞偲峫偊傜傟偰偄傞偨傔丆TAK1偺億儕儐價僉僠儞嵔偵MEKK3傗TRAF6摍偺僔僌僫儖暘巕偑寢崌偡傞偙偲偵傛傝丆暋崌懱偑埨掕壔偟丆僔僌僫儖偑憹嫮偝傟傞婡峔傪峫偊偰偄傑偡丏傑偨丆RING宱楬偐傜Zinc宱楬傊偺僗僀僢僠儞僌丒僔僗僥儉偺夝柧傕廳梫壽戣偺侾偮偱偡丏
丂IL-1僔僌僫儖偼墛徢斀墳傗姶愼杊屼側偳偺柶塽斀墳丆偝傜偵偼娻偺埆惈壔偵傕婑梌偡傞偙偲偑抦傜傟偰偍傝丆堛椕傊偺墳梡偺柺偱傕戝帠側僔僌僫儖偱偡丏偙偙偱婰偟偨傛偆偵IL-1僔僌僫儖偵偍偄偰TRAF6偼尞偲側傞暘巕偱偁傞偨傔丆TRAF6偵傛傞僔僌僫儖揱払婡峔傪偝傜偵徻嵶偵夝柧偡傞偙偲偱丆偦偺抦尒傪婎斦偲偟偨堛椕傊偺墳梡偑婜懸偝傟傑偡丏
