|
|
| 1. 細胞接着分子の異常とがんの浸潤、転移の研究 |
1) CADM1から見る細胞接着研究の面白さ
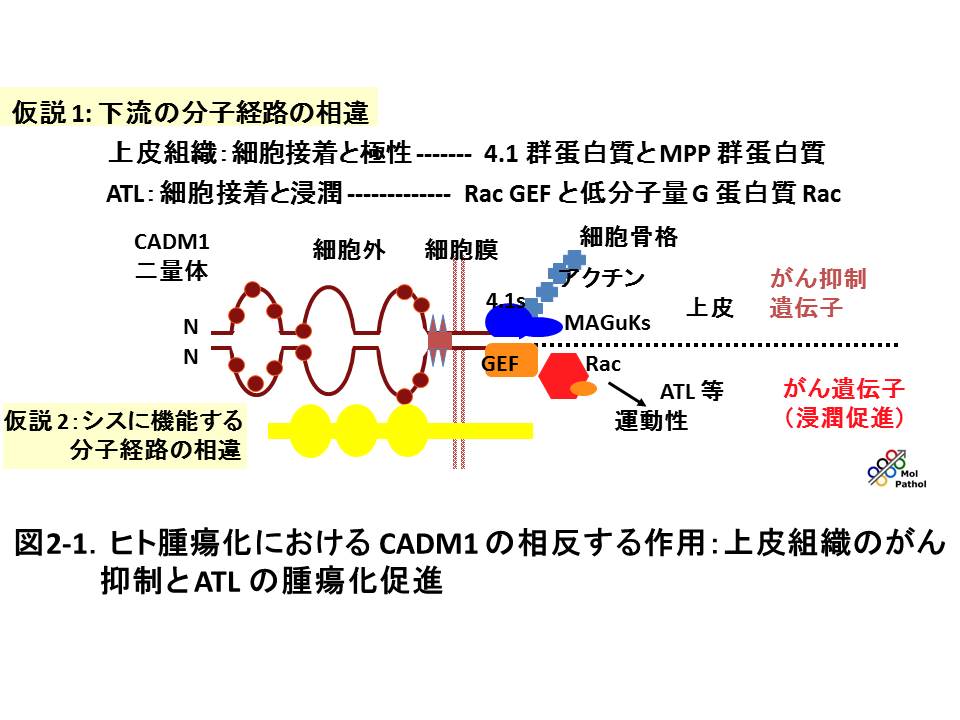
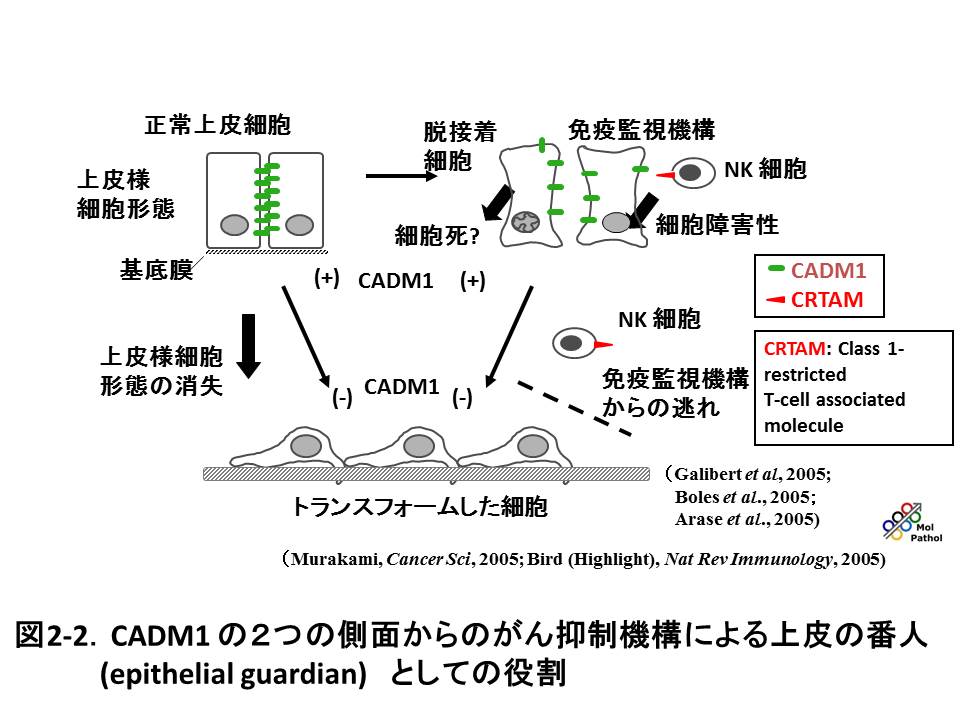
固形がんの死因の90% 以上は浸潤、転移によるものですが、その出発点は、がん細胞同士の接着が失われることです。我々が同定した細胞接着分子CADM1は、免疫グロブリン・スーパーファミリー細胞接着分子をコードし、様々な上皮由来のがん(癌腫)では、がん抑制遺伝子として機能し、ヒトのがん組織、特に進行したがんでは、その発現の欠如や低下が高い頻度で認められます。一方、成人T細胞性白血病(ATL)や小細胞肺癌では特徴的な過剰発現を示し、がん遺伝子として働きます。CADM1が、組織の違いによって、がん化に対し全く正反対の働きを示すことは、例えば、細胞周期の促進分子や阻害分子と比較すると、極めてユニークである反面、複雑でわかりにくい印象を与えるかも知れません。ですがこの意外さ、複雑さにこそ研究の面白さがあるわけですし、実際のがんという疾患は、このような複雑さが幾重にも重なったものだと思います。我々の研究により、CADM1のこの機能の違いは、その直接の下流分子経路や、クロストークする分子経路の違いに起因することがわかってきました。つまり、細胞接着分子は細胞の入口のドアのようなもので、その後ろや傍らにどんな役者が控えているかによって、細胞全体に対する働きが全く変わってしまうということが明らかになりました(図2-1)。さらに、CADM1は上皮細胞間の接着分子としてのみではなく、がん細胞がナチュラルキラー細胞によって認識される際のがん抗原としても働くことが、他のグループの研究により示されました。つまり、CADM1は上皮細胞の接着による正常性を直接保つと同時に、もし上皮細胞の配列が大いに乱れた場合には、免疫監視機構により異常細胞を排除し、結果として組織、個体を守るという二重の意味でがんを抑制していると考えられます。このような二重の機能が示されたがん抑制遺伝子は、CADM1のみです。まさに上皮の番人と言えると我々は思っています(図2-2)。さらに、これらの研究は、がんの浸潤、転移を抑制する医薬品の開発につながると期待されます。現在の分子標的治療薬は、細胞増殖を直接阻害する薬剤がほとんどで、浸潤、転移を抑える医薬品は、様々な理由からまだ開発されていません。我々は、がんの浸潤、転移を制御する全く新しい診断、治療法の開発に結び付く基礎研究を、同時に進めています。
  | |
2) 上皮組織における CADM1のがん抑制遺伝子としての働き
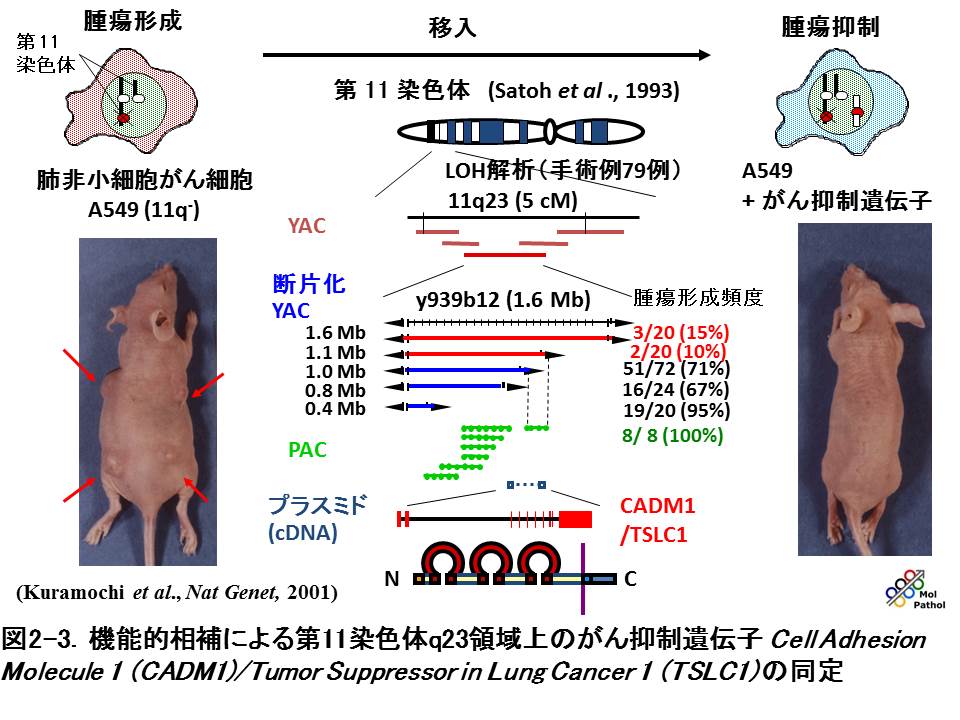
CADM1は、非小細胞肺癌で高頻度に欠失する第11染色体長腕23領域のがん抑制遺伝子として同定したものです。肺がんには、典型的な家族性腫瘍がほとんど見られないことから、他の主だったがん抑制遺伝子が単離されたような連鎖解析によるゲノム構造解析による手法が使えませんでした。そこで、培養肺がん細胞A549に第11染色体長腕23領域のDNA断片を導入し、ヌードマウス皮下での腫瘍形成能の抑制を指標として領域を絞り同定に至りました。この時は、1メガ塩基対(MB)に及ぶヒトゲノムDNA断片をクローニングした酵母人工染色体(YAC)を、スフェロプラスト融合法により
A549細胞へ移入するという方法を取りました。YACは通常、1コピーしかヒトの染色体に組み込まれないので、断片上の遺伝子は内在性のプロモーターによる1コピー分の発現のみが期待されます。CADM1はこのシステムで腫瘍形成能を抑制したことから、その活性が十分強いことが示唆されました(図2-3)。
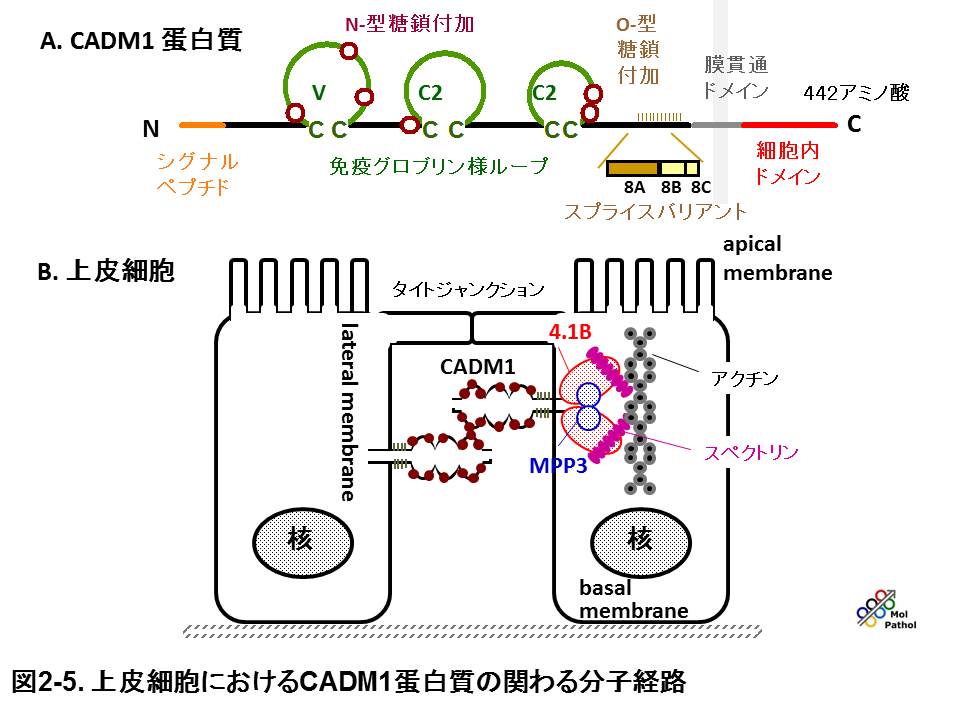
こうして同定したCADM1は、免疫グロブリン・スーパーファミリーに属する膜糖蛋白質をコードし、大部分の上皮組織や神経組織で接着分子として作用します。分化した上皮細胞では、CADM1はホモ2量体を形成して細胞のラテラル面に瀰漫性に発現し、隣接細胞のCADM1ホモ2量体と、トランス結合を形成して、細胞接着に関わると考えられます。さらに我々は、CADM1が細胞内領域の、FERM結合モチーフを介してアクチン結合蛋白質4.1B/DAL-1と、またC末のPDZドメイン結合モチーフを介して裏打ちタンパク質MPP3とそれぞれ結合して、細胞骨格や細胞極性の制御に関わることを示しました(図2-4,
2-5)。さらに、CADM1はsiRNAによってその発現を抑制すると上皮様細胞形態が失われること、また増殖因子受容体METと相互作用をして、HGFによるイヌ腎細胞MDCKの上皮間葉転換(EMT)を実験的に抑えることから、上皮様細胞形態、形質を保持する作用があることがわかりました。
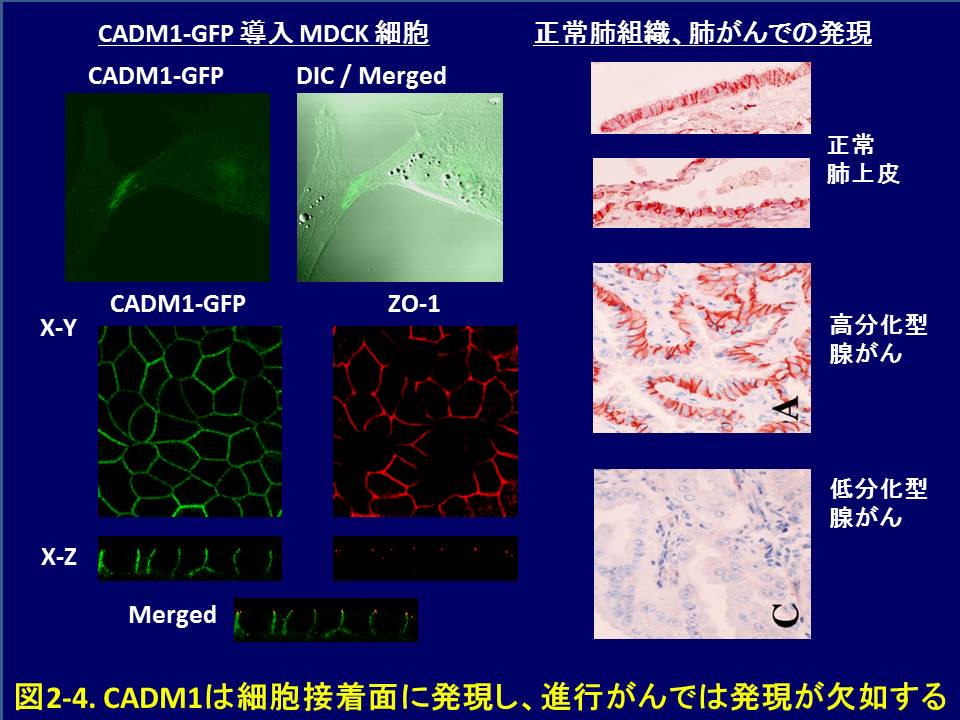
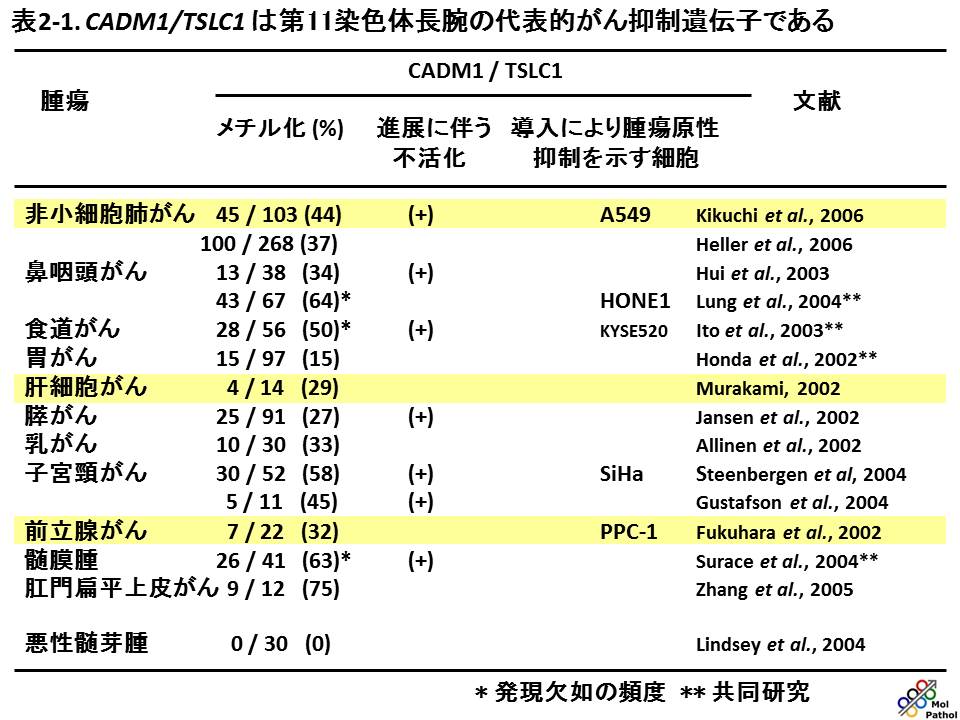
ヒトのがんでは、表2-1に示すように、非小細胞肺がん、乳がん、膵がんなど多くの腫瘍で、プロモーター領域のメチル化などによる2ヒットの不活化が認められ、また発現欠如が認められます。現在ではCADM1は第11染色体長腕上の代表的がん抑制遺伝子とみなされています。非小細胞肺がん組織でCADM1の発現を調べると、非小細胞肺がんでは非浸潤性である
Bronchioloalveolar carcinoma では CADM1の発現が保たれていますが、同じ腫瘍の中でも間質への浸潤性を獲得した
Papillary carcinoma ではCADM1の発現が高率に失われ、CADM1が浸潤に伴って不活化することがわかります。一方、様々な培養がん細胞にCADM1の発現を回復させると、ヌードマウス皮下での腫瘍形成能や、脾臓から肝臓への転移能が抑制され、またCADM1を発現するウイルスベクターをマウスの腫瘍内に注入すると、腫瘍の増殖が抑えられることも示されました。これらの結果は、CADM1が多くのがんで、がん抑制遺伝子として働くことを示しています。
現在は、CADM1が、具体的にどのようにして腫瘍形成を抑制するのかについて、増殖因子受容体経路とのクロストーク、細胞脱接着によるアポトーシスの誘導、CADM1の糖鎖修飾による機能修飾、細胞膜上での安定性の変化などの分子機構を想定して解明を進め、多くの興味深い結果を得ています。CADM1の糖鎖構造の解析は、島津製作所田中耕一記念質量分析研究所の田中耕一フェローのグループとの共同研究です。現在進行中の研究の詳しい内容については、ラボのメンバーに加わった方に御紹介したいと思います。
|
|
3) ATL における CADM1のがん遺伝子(浸潤促進因子)としての働き
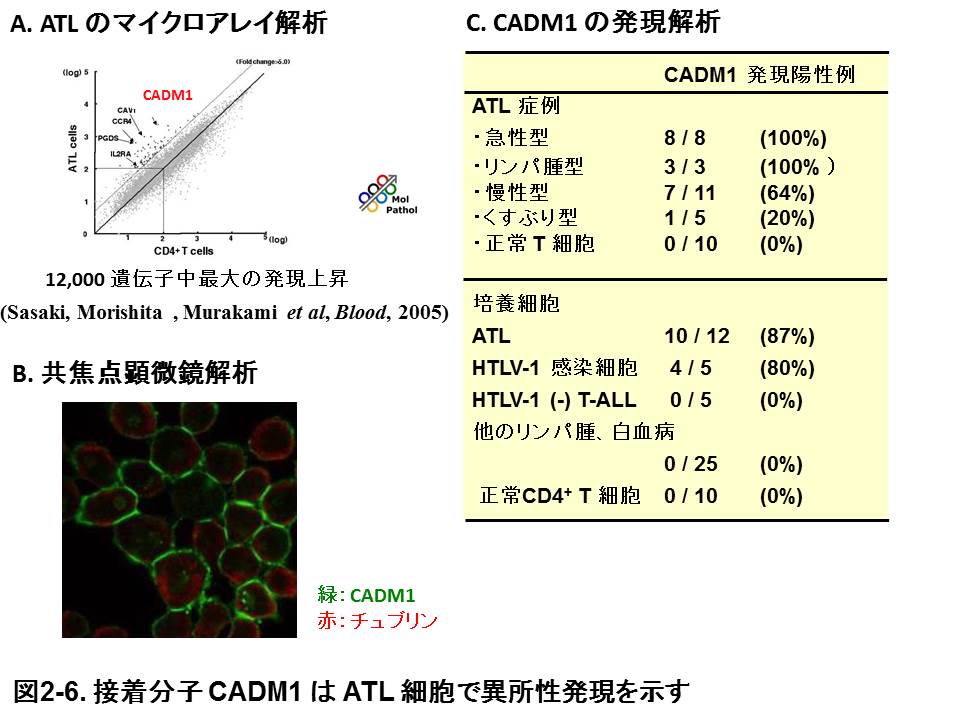
CADM1は多くの上皮組織で発現し、上皮由来の多くのがんでがん抑制遺伝子として働くが、正常リンパ球では全く発現を認めません。ところが、レトロウイルスHTLV-1の感染が引き金となって発症する成人T細胞白血病(ATL)における12,000遺伝子のマイクロアレイ解析から、正常CD4+リンパ球と比較して、急性型ATL細胞で最も発現が上昇する遺伝子としてCADM1が同定されました(図2-6)。CADM1は急性ATL, リンパ腫性ATLで100%、慢性ATL、くすぶり型ATLの一部、さらには無症候性HTLV-1キャリアでもその一部に発現が認められ、悪性ATL細胞の診断マーカーとなることが示されつつあります。従って、CADM1は、上皮とは全く異なり、ATLではがん遺伝子として働く可能性が示唆されます。ATLは他の白血病・リンパ腫と比較して、皮膚や肝臓などの臓器へ強い浸潤性を示すことが特徴です。そこで我々は、ATLにおけるCADM1の発現が、ATLに特徴的な臓器浸潤に関与する可能性を考えて、ATL細胞におけるCADM1下流経路を解析し、ATL細胞では、CADM1はそのC端のPDZ結合モチーフを介して、Tiam1という低分子量Gタンパク質RACのGEFと結合し、RACを活性化することにより、葉状仮足形成を促進し、細胞運動性、浸潤性を高める可能性を示しました。
|
|
4) まとめと今後の課題
上皮とATLにおけるCADM1の相反する機能と分子経路を図2-1にまとめました。CADM1は上皮では、細胞内で 4.1 タンパク質群、MAGuKタンパク質群と結合して細胞骨格制御や上皮様形態の維持に働き、がん抑制遺伝子として機能します。一方、細胞内でTiam1 と結合し、RACを活性化することにより葉状仮足の形成や、細胞の運動性、浸潤性を促進し、ATLでは浸潤促進因子(がん遺伝子)として働きます。下流経路以外にも、我々は最近、細胞膜上の脂質ラフトで、他の増殖因子受容体と複合体を形成して、その働きを修飾することも見出し、研究を続けています。このように、細胞コンテクスト依存的に特定の分子が機能を変えることは、CADM1以外にも、幾つか報告されるようになりました。また、細胞接着分子が増殖因子受容体と複合体を形成して、そのシグナルを修飾することも、他でも報告はありますが、まだ新しく、非常に興味深い領域です。
|
|
次へ
もどる
|
|
|