Division of Cellular and Molecular Biology
The Institute of Medical Science
The University of Tokyo
The Institute of Medical Science
The University of Tokyo
TRAFの話 第二回 -TRAF6と自己免疫病ー
ここで述べる研究は,准教授の秋山泰身先生が中心になって遂行し2005年にScienceに掲載されたものです。
Akiyama, T., Maeda, S., Yamane, S., Ogino, K., Kasai, M., Kajiura, F., Matsumoto, M., and Inoue, J. Dependence of Self-tolerance on TRAF6-directed Development of Thymic Stroma. Science 308, 248-251, (2005))
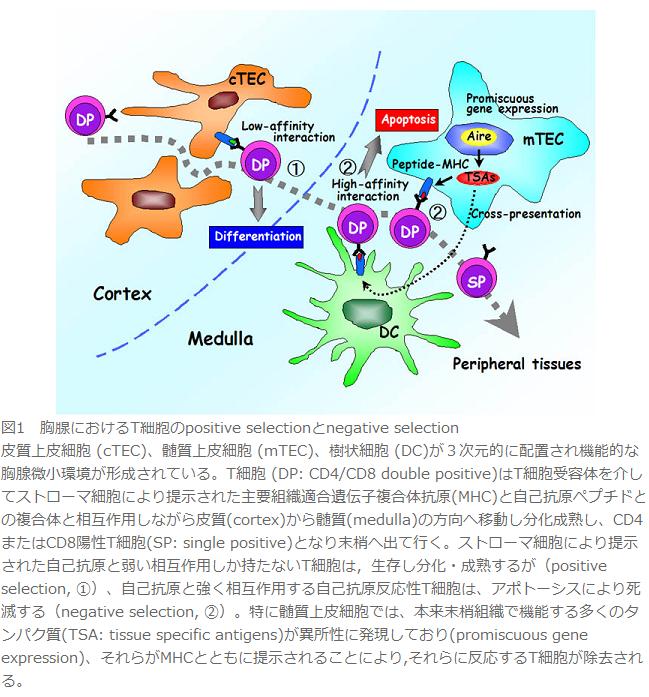
図1 胸腺におけるT細胞のpositive selectionとnegative selection
皮質上皮細胞 (cTEC)、髄質上皮細胞 (mTEC)、樹状細胞 (DC)が3次元的に配置され機能的な胸腺微小環境が形成されている。T細胞 (DP: CD4/CD8 double positive)はT細胞受容体を介してストローマ細胞により提示された主要組織適合遺伝子複合体抗原(MHC)と自己抗原ペプチドとの複合体と相互作用しながら皮質(cortex)から髄質(medulla)の方向へ移動し分化成熟し、CD4またはCD8陽性T細胞(SP: single positive)となり末梢へ出て行く。ストローマ細胞により提示された自己抗原と弱い相互作用しか持たないT細胞は,生存し分化・成熟するが(positive selection, ①)、自己抗原と強く相互作用する自己抗原反応性T細胞は、アポトーシスにより死滅する(negative selection, ②)。特に髄質上皮細胞では、本来末梢組織で機能する多くのタンパク質(TSA: tissue specific antigens)が異所性に発現しており(promiscuous gene expression)、それらがMHCとともに提示されることにより,それらに反応するT細胞が除去される。
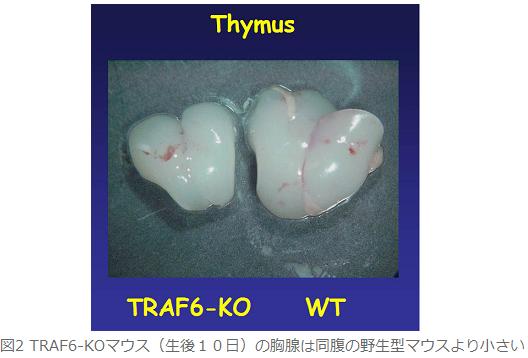
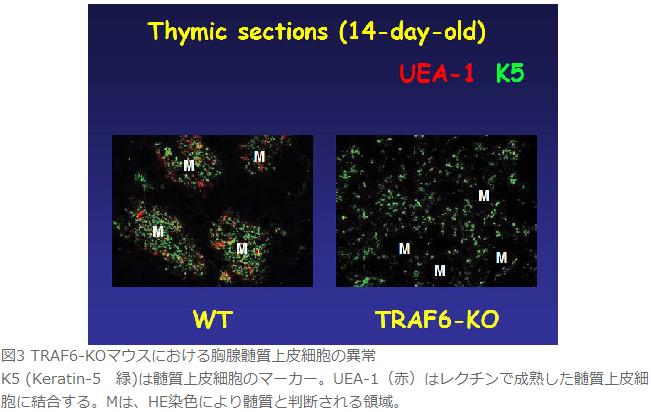
何年も前からTRAF6欠損マウスの胸腺が野生型に比べて萎縮していることに気付いていた(図2)。そこで胸腺のパラフィン組織切片を作製し、ヘマトキシリンーエオシン(HE)染色により組織学的に解析した。その結果、TRAF6欠損マウスでは髄質部分が萎縮していた。また髄質と皮質の境界領域が不明瞭であった。さらに胸腺ストローマを免疫組織染色により検討した結果、TRAF6欠損マウスの胸腺ではUEA-1結合性髄質上皮細胞(成熟髄質上皮細胞)がほとんど存在していなかった。また髄質上皮細胞のマーカーであるKeratin-5の発現を調べたところ野生型では胸腺髄質に局在するが、TRAF6欠損マウスの胸腺では胸腺全体に散在していた(図3)。一方、皮質上皮細胞や樹状細胞の分布は野生型と比べて顕著な差がなかった。以上の結果はTRAF6によるシグナルが髄質上皮細胞の分化成熟とその胸腺内における空間的配置をコントロールしていることが明らかとなった。
胸腺におけるnegative selectionは主として胸腺内に存在する樹状細胞が担っていると考えられてきた。樹状細胞がnegative selectionを行うためには、自己抗原を提示する必要がある。自己抗原が樹状細胞内でも発現している普遍的なタンパク質であれば問題は少ないが、組織特異的抗原(胸腺以外の末梢の臓器や組織で発現している自己抗原)の場合、樹状細胞は組織から血中などに流出してくるタンパク質を貪食する必要があり、効率が悪そうである。前述のようにKyewskiらのグループは胸腺髄質上皮細胞が組織特異的抗原を異所性に発現することでそれらを認識するT細胞のnegative selectionに関与していることを提唱した(図1)。一方、Autoimmune regulator (Aire)はヒト自己免疫疾患APECED (autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy: 自己免疫性多腺性内分泌不全症-カンジダ症-外胚葉ジストロフィー)の原因遺伝子aireの産物である。Aireは胸腺髄質上皮細胞で高く発現しており、Aireの不活化は髄質上細胞における組織特異的抗原の異所性発現を抑制する。即ちAireは胸腺髄質上皮細胞での組織特異的抗原の発現や抗原提示を制御することで胸腺内のnegative selectionに必須の働きをしていると考えられている(図1)。TRAF6欠損マウス胸腺内でのAireの発現および組織特異的抗原の発現を検討した結果、いずれも野生型に比べて著しく減少していた。
ではTRAF6は胸腺髄質上皮細胞においてどのように機能しているのであろうか。その問に答えるため、TRAF6のシグナルに関連する分子を中心に、同様の表現型を示す遺伝子変異マウスを探したところ、RelB欠損マウスに行き当たった。RelBはRel/NF-κBファミリーに属する転写因子であり、その欠損マウスはTRAF6欠損マウスと同様に様々な臓器で自己反応性T細胞が原因の炎症性細胞浸潤が観察される。TRAF6欠損マウスとRelB欠損マウスの表現型は胸腺微小環境形成の異常、自己免疫疾患様の病態の点でほぼ同じであるため、TRAF6とRelBの間に機能的な相関を予想した。そこでTRAF6欠損マウスの胸腺ストローマでのRelBの発現を調べた結果、著しく減少していることが判明した。すなわちTRAF6は胸腺髄質上皮細胞においてRelBの活性制御ではなくRelBの発現そのものを制御している可能性が高い。そこで胎仔胸腺よりTRAF6欠損胸腺髄質上皮細胞を樹立し、RelBの発現を確認したところ、予想通りRelBの発現は著しく減少していた。またそれにTRAF6を再導入したところRelBの発現上昇が起きた。以上の結果はTRAF6が胸腺髄質上皮細胞内でRelB発現量を正に制御していることを強く示唆する。さらにヒトおよびマウスゲノム内でRelB遺伝子のプロモーター領域にはNF-κBとAP-1の結合サイトが存在しており、TRAF6はNF-κBやAP-1を活性化することでRelB遺伝子の発現を誘導すると考察できる。
aly/alyマウスはNF-κB inducing kinase (NIK)遺伝子に機能欠損型の点変異を有する自然発生マウスである。aly/alyもTRAF6欠損マウスやRelB欠損マウスと同様に胸腺髄質上皮細胞の異常や様々な臓器における炎症性細胞浸潤を持つ。徳島大学の松本満先生のグループはaly/alyマウスの胎仔胸腺ストローマをヌードマウスに移植することでNIKが胸腺ストローマ依存的な自己免疫抑制に必須であることを証明した。NIKはTNFRスーパーファミリーから伝達されるNF-κB活性化シグナル経路の中で、いわゆる非古典的経路の活性化に必須である。NIKの活性化はIKKαを活性化し、ついでp100のリン酸化、さらにp100からp52へのプロセシング反応を誘導する。そしてp52/RelB複合体が核移行し転写を活性化する。NIK欠損マウスの胸腺ストローマでは、TRAF6欠損マウスと同様にRelBの発現が減少していると伴にp100からp52へのプロセシングが阻害されている。ところがTRAF6欠損胸腺髄質上皮細胞ではp100からp52へのプロセシングはほとんど変化がなかった。この事実はTRAF6が単純にNIKの上流に位置するのではなく、別の経路によりNF-κBの活性化、RelBの発現誘導を行うことを示唆する。
ところで、LTβR(リンホトキシンβレセプタ-)はTNFRスーパーファミリーの一つであるが、その欠損マウスでは、リンパ節及びパイエル板の欠損の他に肺、肝臓などの臓器に炎症性細胞浸潤が見られる。LTβR欠損マウスは胸腺構造に目立った表現型はみられない。。一方でLTβR欠損マウスでは胸腺内でのAireと組織特異的抗原の発現が減少している。さらにアゴニスティックな抗LTβR抗体をマウスに静脈注射すると胸腺内のAireや組織特異的抗原の発現が上昇するとの報告がある。以上の結果はLTβRからのシグナルは胸腺髄質上皮細胞の分化そのものには必須ではないが、Aireの発現およびそれによる組織特異的抗原の発現と抗原提示に必要であると考察される。
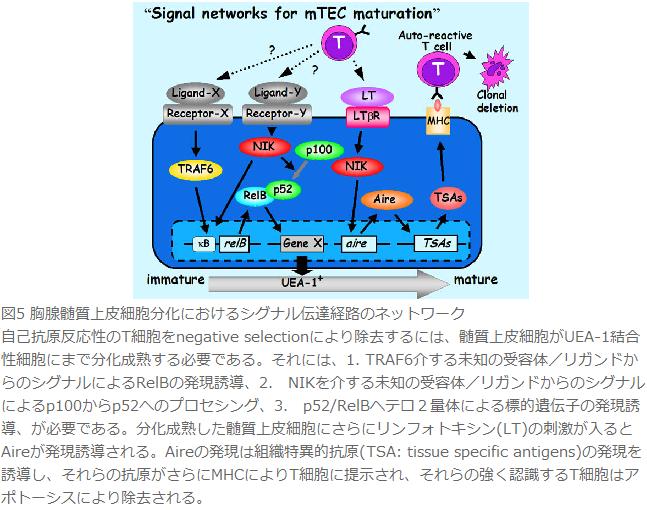
ここまで実験結果をもとにTRAF6、NIK、RelB、LTβRが胸腺髄質上皮細胞の分化とそれによるnegative selectionにどのように関わっているのか考察してみたい(図5)。TRAF6はNF-κBの古典的経路による活性化やAP-1の活性化を介してRelBの発現を誘導する。RelBはNIKによるNF-κBの非古典的経路で誘導されたp100のプロセシング産物p52とヘテロ2量体を形成し、胸腺髄質上皮細胞の成熟に必要な遺伝子転写を行う。この際、TRAF6を介した転写の活性化による遺伝子発現も必要であろう。ついで分化した髄質上皮細胞上のLTβRが刺激を受けることによりNIKによるNF-κBの非古典的経路が活性化されAireや組織特異的抗原の発現が誘導されると考えられる。
現段階では夢物語であるが、免疫機構において自己と非自己の選択性を人為的に制御できれば、移植免疫、自己免疫疾患治療、癌免疫療法など様々な応用が期待できる。ここで記したように免疫系が自己と非自己を識別する機構を解明する上で、TRAF6は鍵となる分子の一つである。今後、TRAF6やそのシグナルの関連分子を明らかにし、それらを標的とした医療への応用が期待される。
Akiyama, T., Maeda, S., Yamane, S., Ogino, K., Kasai, M., Kajiura, F., Matsumoto, M., and Inoue, J. Dependence of Self-tolerance on TRAF6-directed Development of Thymic Stroma. Science 308, 248-251, (2005))
- T細胞は、胸腺の微小環境で自己-非自己の区別を学ぶ
図1 胸腺におけるT細胞のpositive selectionとnegative selection
皮質上皮細胞 (cTEC)、髄質上皮細胞 (mTEC)、樹状細胞 (DC)が3次元的に配置され機能的な胸腺微小環境が形成されている。T細胞 (DP: CD4/CD8 double positive)はT細胞受容体を介してストローマ細胞により提示された主要組織適合遺伝子複合体抗原(MHC)と自己抗原ペプチドとの複合体と相互作用しながら皮質(cortex)から髄質(medulla)の方向へ移動し分化成熟し、CD4またはCD8陽性T細胞(SP: single positive)となり末梢へ出て行く。ストローマ細胞により提示された自己抗原と弱い相互作用しか持たないT細胞は,生存し分化・成熟するが(positive selection, ①)、自己抗原と強く相互作用する自己抗原反応性T細胞は、アポトーシスにより死滅する(negative selection, ②)。特に髄質上皮細胞では、本来末梢組織で機能する多くのタンパク質(TSA: tissue specific antigens)が異所性に発現しており(promiscuous gene expression)、それらがMHCとともに提示されることにより,それらに反応するT細胞が除去される。
- TRAF6は胸腺の機能的な微小環境形成に必須である
何年も前からTRAF6欠損マウスの胸腺が野生型に比べて萎縮していることに気付いていた(図2)。そこで胸腺のパラフィン組織切片を作製し、ヘマトキシリンーエオシン(HE)染色により組織学的に解析した。その結果、TRAF6欠損マウスでは髄質部分が萎縮していた。また髄質と皮質の境界領域が不明瞭であった。さらに胸腺ストローマを免疫組織染色により検討した結果、TRAF6欠損マウスの胸腺ではUEA-1結合性髄質上皮細胞(成熟髄質上皮細胞)がほとんど存在していなかった。また髄質上皮細胞のマーカーであるKeratin-5の発現を調べたところ野生型では胸腺髄質に局在するが、TRAF6欠損マウスの胸腺では胸腺全体に散在していた(図3)。一方、皮質上皮細胞や樹状細胞の分布は野生型と比べて顕著な差がなかった。以上の結果はTRAF6によるシグナルが髄質上皮細胞の分化成熟とその胸腺内における空間的配置をコントロールしていることが明らかとなった。
胸腺におけるnegative selectionは主として胸腺内に存在する樹状細胞が担っていると考えられてきた。樹状細胞がnegative selectionを行うためには、自己抗原を提示する必要がある。自己抗原が樹状細胞内でも発現している普遍的なタンパク質であれば問題は少ないが、組織特異的抗原(胸腺以外の末梢の臓器や組織で発現している自己抗原)の場合、樹状細胞は組織から血中などに流出してくるタンパク質を貪食する必要があり、効率が悪そうである。前述のようにKyewskiらのグループは胸腺髄質上皮細胞が組織特異的抗原を異所性に発現することでそれらを認識するT細胞のnegative selectionに関与していることを提唱した(図1)。一方、Autoimmune regulator (Aire)はヒト自己免疫疾患APECED (autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy: 自己免疫性多腺性内分泌不全症-カンジダ症-外胚葉ジストロフィー)の原因遺伝子aireの産物である。Aireは胸腺髄質上皮細胞で高く発現しており、Aireの不活化は髄質上細胞における組織特異的抗原の異所性発現を抑制する。即ちAireは胸腺髄質上皮細胞での組織特異的抗原の発現や抗原提示を制御することで胸腺内のnegative selectionに必須の働きをしていると考えられている(図1)。TRAF6欠損マウス胸腺内でのAireの発現および組織特異的抗原の発現を検討した結果、いずれも野生型に比べて著しく減少していた。
- 胸腺髄質上皮細胞に必要なシグナル伝達
ではTRAF6は胸腺髄質上皮細胞においてどのように機能しているのであろうか。その問に答えるため、TRAF6のシグナルに関連する分子を中心に、同様の表現型を示す遺伝子変異マウスを探したところ、RelB欠損マウスに行き当たった。RelBはRel/NF-κBファミリーに属する転写因子であり、その欠損マウスはTRAF6欠損マウスと同様に様々な臓器で自己反応性T細胞が原因の炎症性細胞浸潤が観察される。TRAF6欠損マウスとRelB欠損マウスの表現型は胸腺微小環境形成の異常、自己免疫疾患様の病態の点でほぼ同じであるため、TRAF6とRelBの間に機能的な相関を予想した。そこでTRAF6欠損マウスの胸腺ストローマでのRelBの発現を調べた結果、著しく減少していることが判明した。すなわちTRAF6は胸腺髄質上皮細胞においてRelBの活性制御ではなくRelBの発現そのものを制御している可能性が高い。そこで胎仔胸腺よりTRAF6欠損胸腺髄質上皮細胞を樹立し、RelBの発現を確認したところ、予想通りRelBの発現は著しく減少していた。またそれにTRAF6を再導入したところRelBの発現上昇が起きた。以上の結果はTRAF6が胸腺髄質上皮細胞内でRelB発現量を正に制御していることを強く示唆する。さらにヒトおよびマウスゲノム内でRelB遺伝子のプロモーター領域にはNF-κBとAP-1の結合サイトが存在しており、TRAF6はNF-κBやAP-1を活性化することでRelB遺伝子の発現を誘導すると考察できる。
aly/alyマウスはNF-κB inducing kinase (NIK)遺伝子に機能欠損型の点変異を有する自然発生マウスである。aly/alyもTRAF6欠損マウスやRelB欠損マウスと同様に胸腺髄質上皮細胞の異常や様々な臓器における炎症性細胞浸潤を持つ。徳島大学の松本満先生のグループはaly/alyマウスの胎仔胸腺ストローマをヌードマウスに移植することでNIKが胸腺ストローマ依存的な自己免疫抑制に必須であることを証明した。NIKはTNFRスーパーファミリーから伝達されるNF-κB活性化シグナル経路の中で、いわゆる非古典的経路の活性化に必須である。NIKの活性化はIKKαを活性化し、ついでp100のリン酸化、さらにp100からp52へのプロセシング反応を誘導する。そしてp52/RelB複合体が核移行し転写を活性化する。NIK欠損マウスの胸腺ストローマでは、TRAF6欠損マウスと同様にRelBの発現が減少していると伴にp100からp52へのプロセシングが阻害されている。ところがTRAF6欠損胸腺髄質上皮細胞ではp100からp52へのプロセシングはほとんど変化がなかった。この事実はTRAF6が単純にNIKの上流に位置するのではなく、別の経路によりNF-κBの活性化、RelBの発現誘導を行うことを示唆する。
ところで、LTβR(リンホトキシンβレセプタ-)はTNFRスーパーファミリーの一つであるが、その欠損マウスでは、リンパ節及びパイエル板の欠損の他に肺、肝臓などの臓器に炎症性細胞浸潤が見られる。LTβR欠損マウスは胸腺構造に目立った表現型はみられない。。一方でLTβR欠損マウスでは胸腺内でのAireと組織特異的抗原の発現が減少している。さらにアゴニスティックな抗LTβR抗体をマウスに静脈注射すると胸腺内のAireや組織特異的抗原の発現が上昇するとの報告がある。以上の結果はLTβRからのシグナルは胸腺髄質上皮細胞の分化そのものには必須ではないが、Aireの発現およびそれによる組織特異的抗原の発現と抗原提示に必要であると考察される。
ここまで実験結果をもとにTRAF6、NIK、RelB、LTβRが胸腺髄質上皮細胞の分化とそれによるnegative selectionにどのように関わっているのか考察してみたい(図5)。TRAF6はNF-κBの古典的経路による活性化やAP-1の活性化を介してRelBの発現を誘導する。RelBはNIKによるNF-κBの非古典的経路で誘導されたp100のプロセシング産物p52とヘテロ2量体を形成し、胸腺髄質上皮細胞の成熟に必要な遺伝子転写を行う。この際、TRAF6を介した転写の活性化による遺伝子発現も必要であろう。ついで分化した髄質上皮細胞上のLTβRが刺激を受けることによりNIKによるNF-κBの非古典的経路が活性化されAireや組織特異的抗原の発現が誘導されると考えられる。
- 今後の課題
現段階では夢物語であるが、免疫機構において自己と非自己の選択性を人為的に制御できれば、移植免疫、自己免疫疾患治療、癌免疫療法など様々な応用が期待できる。ここで記したように免疫系が自己と非自己を識別する機構を解明する上で、TRAF6は鍵となる分子の一つである。今後、TRAF6やそのシグナルの関連分子を明らかにし、それらを標的とした医療への応用が期待される。
