Division of Cellular and Molecular Biology
The Institute of Medical Science
The University of Tokyo
The Institute of Medical Science
The University of Tokyo
TRAF偺榖丂戞堦夞
- TRAF偺楌巎
- TRAF偼NFκB偲MAPK傪妶惈壔偡傞傾僟僾僞乕僞儞僷僋幙
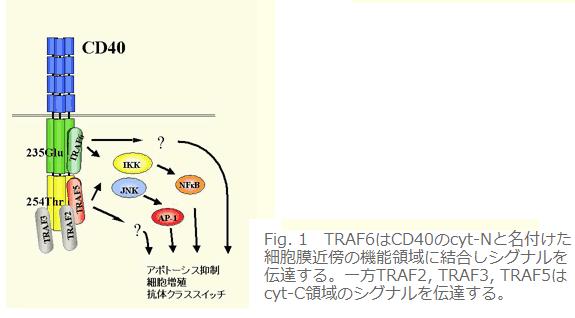
丂
丂TRAF偑悽偵弌傞埲慜偐傜CD40偺僔僌僫儖偑揮幨場巕NFκB傗MAP僉僫乕僛 (JNK, p38, Erk)傪妶惈壔偡傞偙偲偼曬崘偝傟偰偄偨丅偦偙偱摉慠丄偙偺妶惈壔僔僌僫儖傪TRAF偑揱払偡傞偺偱偼偲峫偊偨丅偙傟傪姰慡偵徹柧偡傞偵偼僲僢僋傾僂僩儅僂僗埥偄偼僲僢僋傾僂僩嵶朎偑昁梫偩偐丄偦傟偵偼傑偩帪娫偑昁梫偱偁偭偨丅崟偱偁傞偙偲偼尵偊側偔偰傕尷傝側偔崟偵嬤偄奃怓偲偄偆偙偲傪幚尡揑偵帵偡偙偲偵偟偨丅傑偢丄TRAF5, TRAF6傪攟梴嵶朎偵堦夁惈偵夁忚敪尰偝偣傞偲NFκB, JNK傪妶惈壔偟偨丅偙傟偱惗棟揑忦審偱側偄偵偣傛TRAF5, TRAF6偼偙傟傜偺妶惈壔僔僌僫儖傪揱払壜擻偲偄偆偙偲偵側傞丅師偵CD40偺嵶朎幙椞堟偺偳偙偵TRAF偑寢崌偡傞偐偲偄偆栤戣偵摎偊傞偙偲偵偟偨丅寢嬊TRAF僼傽儈儕乕偺拞偱CD40偵捈愙寢崌偡傞偺偼TRAF2, TRAF3, TRAF5, TRAF6偱偁傝丄偝傜偵偦偺拞偱TRAF2, TRAF3, TRAF5偼枌偐傜墦偄摨偠晹埵乮変尋媶幒偱偼cyt-C偲屇傇丅乯偵寢崌偡傞偑TRAF6偩偗偼枌嬤朤晹埵乮cyt-N乯偵寢崌偟偨丅偝傜偵奺乆偺寢崌偼偨偭偨堦偮偺傾儈僲巁抲姺偱徚幐偟偨丅懄偪TRAF2, TRAF3, TRAF5偺偮偄偰偼254斣栚偺Thr傪Ala偵抲姺乮T254A曄堎乯偡傞丄TRAF6偺応崌偵偼235斣栚偺Glu傪Ala偵抲姺乮E235A曄堎乯偡傞偲慡偔寢崌偟側偔側傞丅偝傜偵戝帠側偙偲偼cyt-N偲cyt-C偼偦傟偧傟撈棫偵NFκB傪妶惈壔偡傞偙偲偑偱偒傞偺偱偁傞偑丄偙傟傜偺揰曄堎傪摫擖偡傞偙偲偵傛傝偦偺妶惈壔擻偑徚幐偡傞偙偲偱偁傞(Fig. 1)丅偨偭偨堦偮偺傾儈僲巁抲姺偵傛傝TRAF偺寢崌偑徚幐偟摨帪偵偦偺晹埵偐傜偺NFκB妶惈壔僔僌僫儖偑徚偊傞偺偱偁傞偐傜尷傝側偔崟偵嬤偄奃怓偲尵偊傞丅TRAF偺僪儈僫儞僩僱僈僥傿僽曄堎懱傪梡偄偨NFκB妶惈壔偺梷惂幚尡傕惉棫偟偨偑丄奃怓偑偝傜偵擹偔側傞傢偗偱偼側偄丅偙偆側偭偨傜TRAF6僲僢僋傾僂僩儅僂僗傪嶌傞偟偐側偄丅
- TRAF6僲僢僋傾僂僩儅僂僗
TRAF6僲僢僋傾僂僩儅僂僗傪嶌偭偨(Fig. 2)丅

嵟懍偱嶌偭偨偮傕傝偩偭偨偑丄暷崙偺僌儖乕僾偵傎傫偺彮偟愭傪挻偝傟偰偟傑偭偨丅偱傕晄巚媍側偙偲偵乮偄傑偩偵側偤側偺偐偼傢偐傜側偄丅乯変乆偑摨掕偟偨偄偔偮偐偺昞尰宆偑斵摍偑敪昞偟偨傕偺偲堎側偭偰偄偨丅傑偢変乆偑摨掕偟偨昞尰宆傪傑偲傔偰傒傞丅
丒攋崪嵶朎宍惉晄慡偵傛傞崪検偺堎忢憹戝偱崪戝棟愇昦傪敪徢(Fig. 3)丅
丒儕儞僷愡宍惉晄慡(Fig. 4)丅
丒枹惉弉B嵶朎偺暘壔梷惂丅
丒恄宱娗暵娗晄慡丅
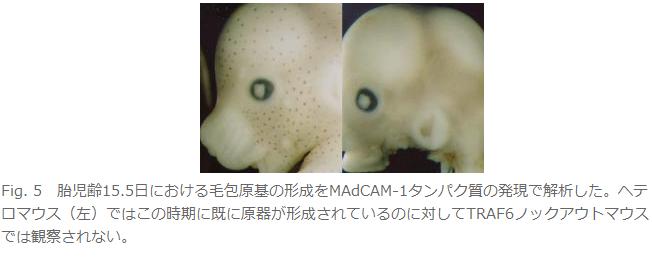
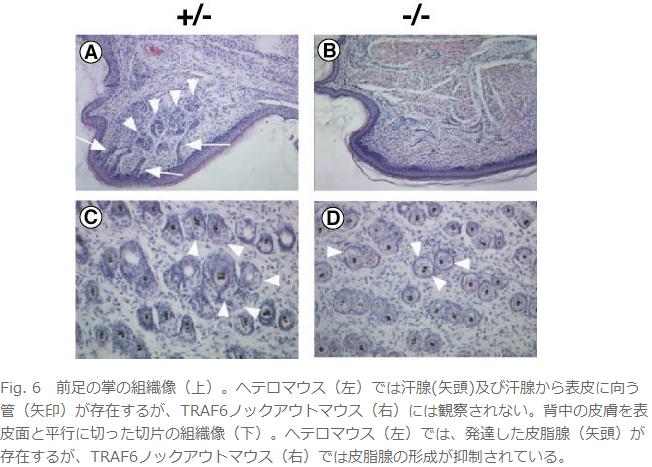
丒栄曪丄娋態丄旂帀態側偳旂晢晅懏婍姱偺宍惉晄慡傪庡徢忬偲偡傞柍娋惈奜泱梩宍惉晄慡徢偺敪丂徢(Fig. 5偲Fig. 6)丅
丒IL-1, LPS僔僌僫儖偺徚幐
丒銪憻偵偍偗傞CD40梲惈B嵶朎偺尭彮丅
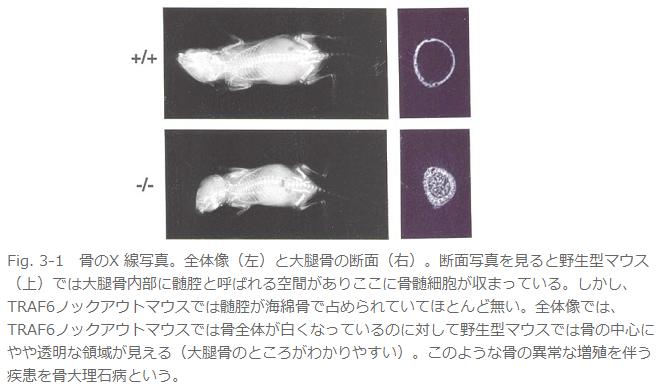
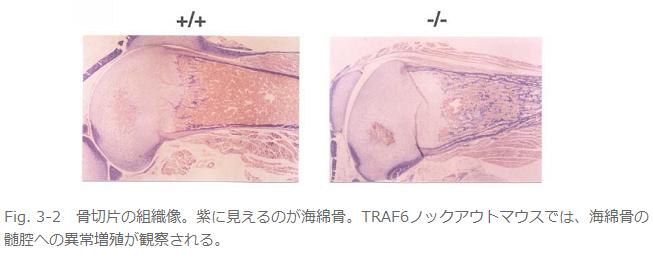
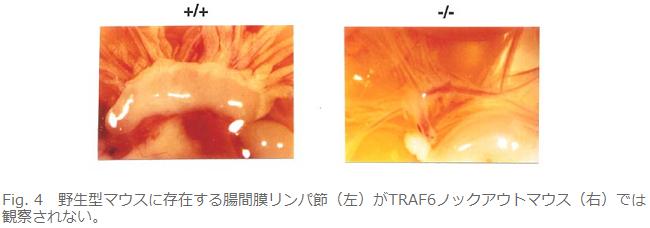
丂偙偺儅僂僗偼堎忢偑懡偄丅傑偢暷崙偺僌儖乕僾傕崪戝棟愇昦偱偁傞偲弎傋偰偄傞丅偦偙傑偱偼摨偠偱偁傞丅偱傕偦偺尨場偑堘偭偰偄偨丅崪戝棟愇昦偺懡偔偼崪傪暘夝偡傞攋崪嵶朎偺婡擻晄慡乮攋崪嵶朎條嵶朎乵亖懡妀偱摿庩側愼怓朄偱愒偔愼傑傞丅乶偑偁偭偰傕偦偺嵶朎偵崪傪暘夝偡傞擻椡偑柍偄丅乯偐丄攋崪嵶朎條嵶朎偝偊柍偄偐偱偁傞丅変乆偼偄偔傜扵偟偰傕攋崪嵶朎傪尒偮偗傜傟側偐偭偨偑丄暷崙僌儖乕僾偼傎傏惓忢側悢偺攋崪嵶朎偑懚嵼偡傞傕偺偺崪媧廂擻偑側偄偲愢柧偟偰偄傞丅曗懌偡傞偲攋崪嵶朎宍惉偵偼慜嬱嵶朎偑梈崌偟偰崪媧廂擻偺柍偄攋崪嵶朎條嵶朎偵側傞乽暘壔乿偺抜奒偲偝傜偵嵶朎偺宍懺偑崪偵媧拝偡傞傛偆偵曄壔偟偰崪媧廂擻傪妉摼偡傞乽妶惈壔乿偺俀抜奒偑偁傞丅偦傟偱変乆偺TRAF6僲僢僋傾僂僩儅僂僗偼暘壔傕妶惈壔傕婲偙傜側偄丅堦曽偱暷崙僌儖乕僾偺TRAF6僲僢僋傾僂僩儅僂僗偼暘壔偼婲偙傞偑妶惈壔偼婲偙傜側偄偲偄偆傢偗偱偁傞丅変乆偼攋崪嵶朎偺慜嬱嵶朎傪TRAF6僲僢僋傾僂僩儅僂僗偐傜庢傝弌偟攟梴偟偰攋崪嵶朎偵暘壔偝偣偰傒偨偑慡偔暘壔偑桿摫偱偒側偐偭偨丅偟偐偟丄偙偺TRAF6寚懝慜嬱嵶朎偵慡挿偺TRAF6傪堚揱巕摫擖偡傞偲崪媧廂擻傪桳偡傞攋崪嵶朎偑桿摫偝傟丄傑偨TRAF6偺RING僼傿儞僈乕傪寚懝偝偣偨TRAF6傪摫擖偡傞偲崪媧廂擻偺柍偄攋崪嵶朎條嵶朎偵暘壔偡傞偙偲偐傜丄TRAF6偼攋崪嵶朎偺暘壔偲妶惈壔偺椉曽偺僗僥僢僾偱婡擻偟偰偄傞偲峫偊傜傟傞丅儕儞僷愡宍惉晄慡丄枹惉弉B嵶朎偺暘壔梷惂丄柍娋惈奜泱梩宍惉晄慡徢丄CD40梲惈B嵶朎偺尭彮偵偮偄偰偼摿偵婰弎偑柍偄丅恄宱娗暵娗晄慡丄IL-1, LPS僔僌僫儖偺徚幐偵偮偄偰偼摨偠昞尰宆偱偁傞丅偙偺傛偆偵椉僌儖乕僾偺儅僂僗偱偐側傝杮幙揑側昞尰宆偺嵎偑偁傞偑丄慜弎偟偨傛偆偵偦偺棟桼偼傢偐傜側偄丅堘偆偲偡傟偽僞乕僎僢僥傿儞僌儀僋僞乕偺峔憿偑堘偆偖傜偄偱偁傠偆丅
- 偙傟偐傜偺TRAF6
丒TRAF6偺偳偺晹埵偑忋棳偐傜偺僔僌僫儖傪庴偗庢傞偺偵昁梫偱偁傞偐丄傑偨壓棳傊僔僌僫儖揱丂払偡傞偺偵偳偺晹埵偑昁梫偐丠
丒TRAF6偺崅師峔憿偼偳偆側偭偰偄偰丄偦傟偼僔僌僫儖揱払偺帪偵偳偆曄壔偡傞偺偐丠
丒TRAF6偺忋棳偲壓棳偺暘巕偼壗偐丠
丒TRAF6偼僔僌僫儖揱払偺帪偵廋忺偝傟傞偐丠
偙偺傛偆側暘巕惗暔妛揑側栤戣揰偺夝寛偑尰嵼偺尋媶幒偺僥乕儅偵娷傑傟傞丅偙偺懠偵TRAF6偑娭傢傞惗柦尰徾傪傕偲偵堛栻昳奐敪傗嵞惗堛椕偵栶棫偰傞偙偲傪峫偊偰偄傞丅TRAF6僲僢僋傾僂僩儅僂僗偺偲偙傠偱弎傋偨傛偆偵TRAF6寚懝偵傛傝攋崪嵶朎宍惉晄慡丄旂晢晅懏婍姱偺宍惉晄慡丄儕儞僷愡宍惉晄慡丄恄宱娗宍惉晄慡側偳恖偺幘姵偲怺偔娭傢傞堎忢偑惗偢傞偙偲偐傜丄TRAF6偑庬乆偺廳梫側婍姱宍惉偵昁恵偱偁傞偙偲偼柧傜偐偱偁傞丅偟偨偑偭偰暘巕惗暔妛揑側栤戣揰偺夝寛偲丄奺乆偺慻怐偵偍偗傞TRAF6偺摿堎惈偺夝柧偑恑傔偽丄TRAF6偁傞偄偼偦偺廃曈傪僞乕僎僢僩偲偟偨堛栻昳奐敪傕壜擻偲峫偊偰偄傞丅傑偨嵞惗堛椕傊偺墳梡偵偮偄偰傕摿偵崪偲旂晢晅懏婍姱偵徟揰傪峣傝尋媶傪恑傔偰峴偔偮傕傝偱偁傞丅
