Research
消化管は食餌性抗原や腸内細菌・真菌など多種多様な抗原に常に曝されている特殊な器官です。例えば、ヒトの消化管にはおよそ100兆個、500~1000種類もの細菌が常に生息し、宿主である我々と平和的な共生関係を築いています。特に腸管では、これら無数の抗原に対処するため、免疫細胞と上皮細胞が互いに作用しながら病原性微生物を「排除」し、非病原性微生物と「共生」する基盤を形成して、腸管の恒常性を維持しています。一方、腸管内に生息する細菌は、宿主に対し栄養の供給や免疫細胞の分化・増殖、病原性微生物の排除など実に様々な作用を発揮しています。宿主とこれら腸内細菌との共生関係が成立する仕組みは生物学的に大変興味深いだけでなく、腸内細菌叢の恒常性が崩れる(Dysbiosisになる)とクローン病や潰瘍性大腸炎といった炎症性腸疾患、肥満や糖尿病をはじめとする生活習慣病、大腸・肝臓ガン、アレルギー疾患などの発症に深く関わることがこれまでの研究で明らかになっています。そのため、腸管免疫細胞と上皮細胞による腸管恒常性制御システムを理解することは、大変重要な研究課題となっています。
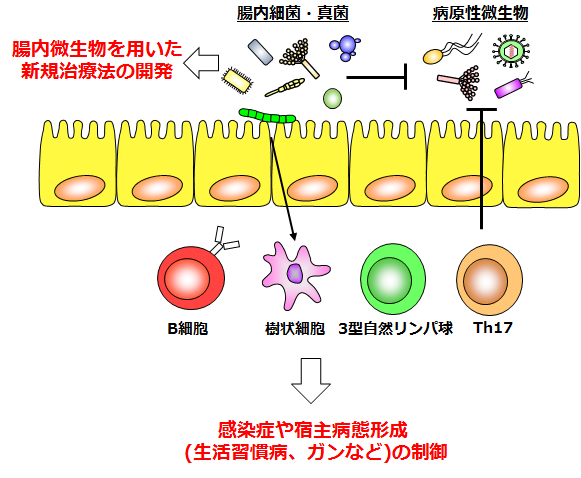
当研究室のミッションは、「腸管における微生物との共生と排除の仕組みを理解し、感染症や腸内微生物が関わる疾患の克服に貢献すること」です。実際、腸内細菌をはじめとする腸管微生物と宿主の研究は、現在、世界中で注目されており、将来新たな研究分野の創生や創薬開発、技術革新が生まれる可能性を秘めています。本研究室では、宿主と腸内細菌間の共生因子であり腸管上皮細胞が発現するα1, 2-フコースの誘導および制御機構を明らかにすることで、腸管恒常性維持システムの解明とその破綻によって引き起こされる様々な疾患の予防・治療・診断法の開発を行っています(腸管上皮細胞のα1,2-フコース誘導・制御機構の解明)。また、腸内細菌は、腸管上皮細胞が発現するα1, 2-フコースだけでなく、T細胞やIgA陽性細胞など腸管免疫細胞の分化誘導、機能付与にも深く関与しています。我々は特に腸管Th17細胞や3型自然リンパ球(Group 3 innate lymphoid cells : ILC3)に着目し、それらの免疫細胞の相互作用や腸管恒常性維持機構の理解を目指して研究を行っています(腸管Th17細胞の恒常性維持機構)。さらに、腸内には細菌だけではなく、真菌やウイルスなど他の微生物も生息しています。これらの微生物が、腸内細菌や宿主とどのように相互作用をしているのか、ほとんど明らかとなっていません。本研究室では特に腸内細菌と真菌に着目し、病原性微生物に対する感染防御機能や宿主病態形成(例:炎症性腸疾患、生活習慣病、ガンなど)にどのように関わっているのか、分子・細胞・個体レベルで解析を進めています(腸内細菌・真菌による宿主病態制御)。
腸管上皮細胞のα1,2-フコース誘導・制御機構の解明
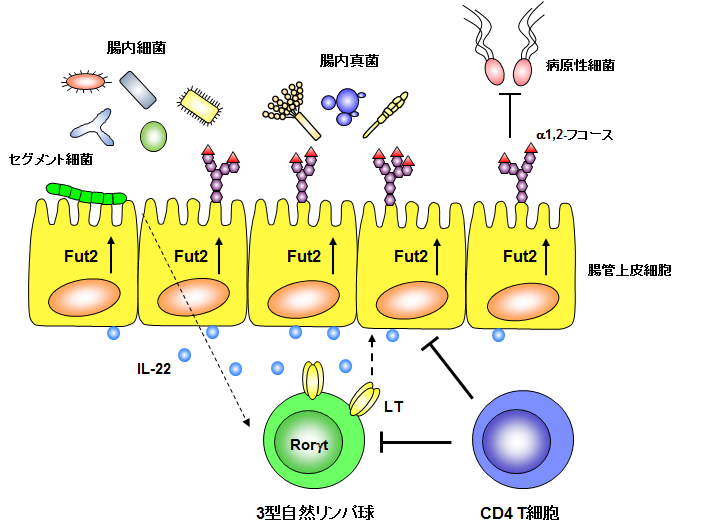
腸管上皮細胞は細胞表面に糖鎖を発現しており、糖鎖の末端にはFucosyltransferase 2 (Fut2)という糖転移酵素によってα1,2-フコースが付加されています。このFut2遺伝子は、ヒトにおいて不活性型の変異多型が報告されており、Fut2不活性型変異多型は、クローン病、I型糖尿病、原発性硬化性胆管炎、乾癬、ベーチェット病の関連遺伝子であることが報告されています。また、腸管上皮細胞が発現するα1,2-フコースは、Bacteroidesなどの腸内細菌に取り込まれて栄養源となることから、宿主と腸内細菌間における共生因子の一つであると考えられています。そのため、腸管上皮細胞のFut2およびα1,2-フコースの発現機構を明らかにすることは、生物学的、医学的に重要な研究課題です。
当研究室では、腸内細菌の一種であるセグメント細菌(segmented filamentous bacteria : SFB)が、腸管における代表的な自然免疫細胞である3型自然リンパ球(Group 3 innate lymphoid cells : ILC3)を刺激することで、IL-22の産生を誘導し、腸管上皮細胞のFut2およびα1,2-フコースの発現を誘導することを見出しました(Goto Y, et al. Science, 345: 1254009 (2014))。一方、獲得免疫細胞であるCD4T細胞がIL-10を産生することで、腸管上皮細胞のFut2およびα1,2-フコースの発現を負に制御することも見出しました(Goto Y, et al. Sci Rep, 5: 15918 (2015))。このことから、1. 腸内細菌と免疫細胞、上皮細胞の3者間ネットワークによって上皮細胞のα1,2-フコースが誘導されること、2. 上皮細胞のα1,2-フコースの発現は、自然免疫細胞(ILC3)と獲得免疫細胞(CD4T細胞)によって調節されることが分かりました(下図)。また、上皮細胞が発現するα1,2-フコースは、病原性細菌の感染を防御することから、これらの結果は腸管における新たな免疫システムの存在を示しています。しかしながら、この誘導・制御機構については未解明の部分が数多く残されています。今後は、腸内細菌と免疫細胞の相互作用や、免疫細胞同士の相互作用について、より詳細な解析を行っていく予定です。また、この上皮細胞のα1,2-フコースを使って、炎症性腸疾患をはじめとする様々な疾患の新たな予防・治療・診断方法の開発にも取り組んでいきます。
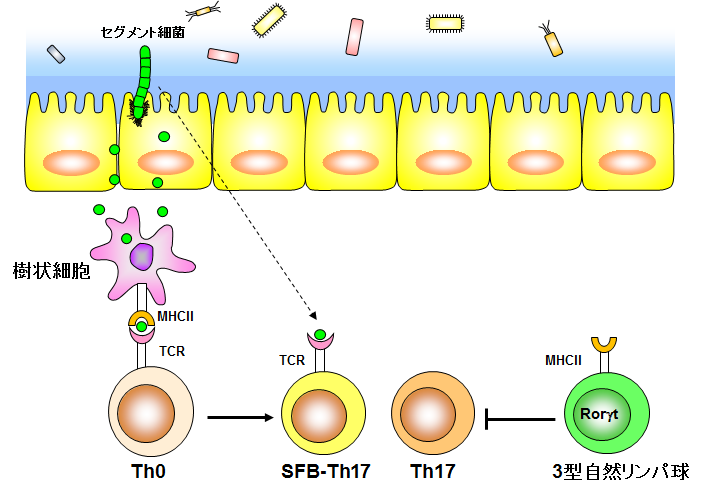
腸管Th17細胞の恒常性維持機構
Th17細胞は、Citrobacter rodentiumやCandida Albicansをはじめとする病原性細菌や真菌の排除に寄与するヘルパーT細胞です。これまでに、腸管Th17細胞は代表的な腸内細菌であるSFBによって誘導され、誘導されたTh17細胞はセグメント細菌由来の抗原を認識し、樹状細胞からの抗原提示を受けて増殖することが報告されています(Goto Y, et al. Immunity, 40: 594-607 (2014))。一方、3型自然リンパ球は細胞表面にMHCII分子を発現し、セグメント細菌非依存的なTh17細胞の分化誘導を負に制御しています(Goto Y, et al. Immunity, 40: 594-607 (2014))。このため、腸管Th17細胞の恒常性は樹状細胞と3型自然リンパ球という2つの自然免疫細胞によって正と負に制御されていることが明らかとなっています(下図)。しかしながら、このセグメント細菌依存的、非依存的に誘導されるTh17細胞の質的な相違や分化様式ついては未だ多くの謎に包まれています。本研究室ではセグメント細菌依存的、非依存的なTh17細胞の性質を明らかにすることで、腸管の恒常性維持や感染防御機構の解明を目指しています。
腸内細菌・真菌による宿主病態制御
ヒトの腸管には無数の細菌が生息しており、病原体の感染防御や宿主の疾患制御に深く関与しています。また、腸内には細菌だけではなく真菌やウイルスなど多数の微生物が存在しています。特に、腸内に生息する真菌がどのような機能を持っているのかについては、まだほとんど研究がなされていません。当研究室ではヒトおよびマウスの腸内細菌・真菌叢に着目し、腸内細菌と真菌の関わりや、腸内細菌・真菌と宿主の相互作用と宿主の病態形成について研究を行っています(下図)。特に、高病原性の感染症や日和見感染症に対する腸内細菌・真菌の防御効果や、炎症性腸疾患、生活習慣病(肥満や糖尿病)、ガンの発症との関わりについて研究を行っています。また、我々は日々の食生活を通して摂取する食餌に含まれる細菌・真菌由来の成分にも着目して研究を行っています。さらに、感染症に対する抑制効果や生活習慣病、ガンの予防・治療効果を持つ腸内細菌・真菌を分離し、それらをターゲットとした新たな治療・予防・診断法(例:マイクロバイオーム、マイコバイオーム創薬)の開発を目指しています。