English version is here

1.腎臓をめぐる現在の状況
腎機能が失われると、水分と種々の毒性物質が蓄積し、意識混濁、肺水腫による呼吸困難、高カリウム血症などで死に至るため、人工透析が必要となる。さらに腎臓は内分泌器官としても重要で、レニンを産生することによって血圧の調節にも関わり、ビタミンDの活性化を通して骨代謝にも関わる。またエリスロポエチンの主要産生臓器であるため、腎不全では赤血球維持に異常をきたし、重度の貧血となる。当研究室をサポートしているAmgen社によるエリスロポエチンの発見によってこの問題は解決されたが、エリスロポエチンはほぼ一生にわたって週に数回投与する必要があり、医療費の高騰を招いている。日本で人工透析を受ける人は20万人を超え、この10年で2倍となった。腎不全は難病指定とされ、その医療費はすべて国庫によって賄われるため、社会的負担は大きい。このような状況にもかかわらず、腎機能が一旦悪化するとそれを改善させる画期的な治療法はいまだ存在せず、最終的には透析導入となる。もし腎臓の幹細胞なるものが存在すれば、あるいはそれを別の細胞源から誘導できれば、それを移植することによって腎機能を改善できないであろうか?それが直ちには難しいとしても、腎臓の発生を探ることによって臓器を再生する手だてが考えられないだろうか。また腎臓発生自体が複雑な臓器形成のモデルとしてchallengingな課題でもあり、長年多くの発生学者を引きつけてきたが、この数年の分子生物学の発展に伴い、分子の言葉で腎臓発生を語れるようになってきた。
2.腎臓の発生
1)腎臓発生の概略
腎臓は中間中胚葉から発生し、前腎、中腎、後腎の3段階を経て形成される。前腎、中腎のほとんどは後に退行変性し、哺乳類成体において機能する腎臓は後腎である。(図1)に示すように、前腎は1つのネフロンからなる非常に単純な構造である。中腎はその尾側に発生し、数十のネフロンからなる。この一部は男性生殖器となるが、腎臓としての中腎は退行する。我々哺乳類の腎臓、すなわち後腎は、中腎管 (ウルフ管)の最も尾側に尿管芽と呼ばれる突起が出現し、その周りに間葉組織が集合して生じる。この尿管芽と後腎間葉との相互作用によって、数百万ものネフロンをもつ後腎が完成する。尿管芽の周りに間葉細胞が凝集し、それが上皮化してS字体といわれる状態を経て、糸球体、近位及び遠位尿細管が発生する(図2)。尿管芽は分岐を重ね、集合管、尿管となる。つまり、腎臓としての機能を司るそのかなりの部分が後腎間葉から発生することになり、尿細管には10種を超える細胞が存在するため、後腎間葉は多能性をもった前駆細胞集団ともいえる。
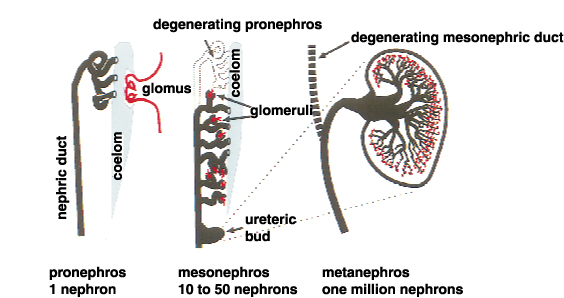
図1.腎臓の構造 |
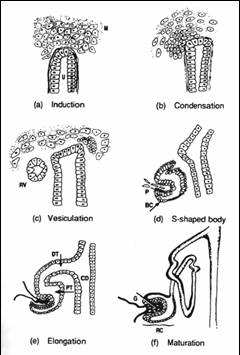
図2.ネフロンの発生 |
2)Znフィンガー蛋白 Sall1の単離
前腎はアフリカツメガエル(Xenopus laevis)で解析が進んでおり、特にアニマルキャップアッセイという予定外胚葉を培養して各種臓器を誘導する系が確立している。東京大学の浅島誠らによって、このアニマルキャップをアクチビンとレチノイン酸の存在下に生理食塩水の中で培養すると、わずか3日で3次元の立体構造をもった前腎管が形成されることが示されている。我々はこの系に着目し、浅島らと共同して、この前腎管を誘導する条件としない条件とで、種々のサブトラクションを行った(図3A)。その中から単離されたのが、Zincフィンガーを8個持つ新規蛋白で、これをXsal-3と名付けた(図3B)。この遺伝子はショウジョウバエのspalt (sal)のホモログで、確かに前腎に発現していた。しかし、その他に中枢神経系、耳胞、鰓弓にも発現しており、前腎特異的とは言えなかった。しかし、この遺伝子を指標にマウス後腎から新たな遺伝子が単離でき、この遺伝子(Sall1)の発現様式を調べたところ、尿管芽が後腎間葉に侵入する胎生11.5日において、尿管芽を取り囲む後腎間葉に非常に強く発現していた(図4)。Sall1は腎臓の他に、中枢神経系、耳胞、心臓、肢芽、肛門などに発現しており、Xsal-3との一部類似性が認められた。さらに、中枢神経系では脳室周囲の神経幹細胞が存在する領域に、肢芽ではprogress zoneという未分化細胞が増殖する部分で発現が認められ、腎臓に限らず他の未分化細胞でも何らかの役割を持つ可能性が示唆された。
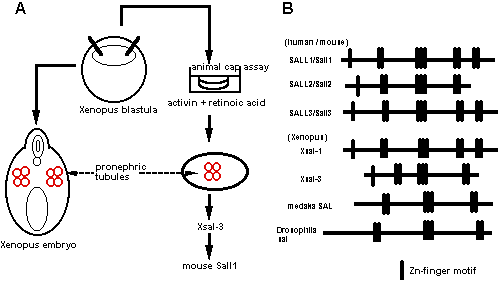
図3.Salの単離 |
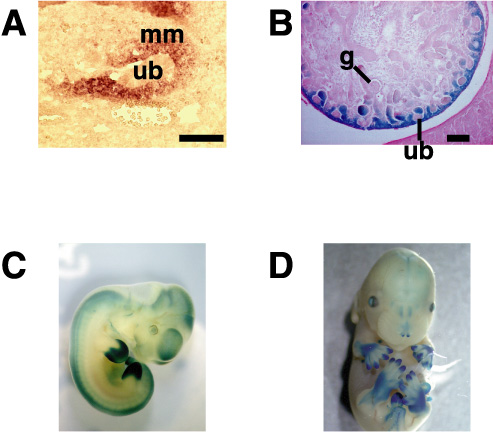
図4.マウスSall1の発現パターン |
3)Sall1は腎臓発生に必須である
そこでSall1を欠失するマウスを作成したところ、すべてのノックアウトマウスが生直後に死亡した。開腹してみると、腎臓が完全に欠損しているか、非常に小さい痕跡的な腎臓が認められるのみであったため(図5)、Sall1は腎臓の発生に必須であることが証明された。さらにノックアウトマウスでは、尿管芽は後腎間葉に侵入していないか、あるいはしてもその後の分岐は著明に障害されていた。つまりSall1が、後腎発生の最も初期段階の重要なステップである尿管芽の侵入に必須であることが判明した。後腎間葉にSall1が発現することによって、尿管芽を引き寄せる機構は一体何か?Sall1が核内でヘテロクロマチンに存在することも合わせて、Sall1の分子メカニズムの解明を急いでいる。
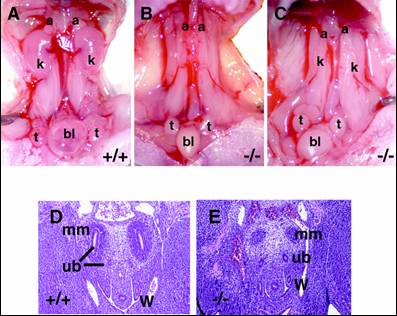
図5.Sall1ノックアウトマウス |
2.後腎間葉に幹細胞はあるのか?
後腎間葉は尿管芽の分岐とともにそれを取り囲むように分布するので、どんどん数が増え一つ一つは小さくなっていく。齧歯類においてはこのような間葉組織は生後数日まで存在するが、成体においては認められない。この間葉組織が失われることが、成体において腎臓が修復しない原因ではないだろうか?とすれば、腎臓の前駆細胞集団である間葉組織を何らかの方法で誘導したり移植したりできれば成体での腎臓修復も可能になるかもしれない。この目的のために、ES細胞から腎臓前駆細胞の誘導法の確立、腎臓前駆細胞のin vitro, in vivo の分化系の確立を試みている。このような腎臓における分化系が確立されれば、あとは様々な細胞表面マーカーでソートすることによって、腎臓前駆細胞を単離していくことができるはずである。そして、これをin vitroで未分化なまま増幅し、必要に応じて移植し腎機能を回復させることが究極の目的となる。
3.ES細胞未分化維持因子の同定
前述した通りSall1は、脳室周囲の神経幹細胞が存在する領域や、progress zoneという肢芽の未分化細胞が増殖する部分など、腎臓に限らず他の未分化細胞でも発現が認められる。またES細胞でも非常に強く発現している。このことから幹細胞に共通の機構の存在が示唆される。これにヒントを得て、現在マウスおよびサルES細胞を未分化状態で維持できる因子を同定している(未発表)。LIFは確かに マウスES細胞を維持する因子ではあるが、大抵のES細胞はさらにfeeder細胞が必要で、このfeederが何を供給しているか判明していない。サルとヒトのESにはfeederは必要だが LIFは効果がないことから、LIF以外の因子の重要性が認識されている。我々の同定した因子がこれに相当するのか、 Oct3/4やLIF/STAT3とどのような関係にあるのか、ヒトES細胞には効果があるのか、といった問題をこれから検討していきたい。
4.おわりに
腎臓発生・再生をSall1を軸として解明していくことが主目的であるが、ES、マウス、カエルを並行して使っているため、得られた知見を直ちに違う系に投入することが可能である。腎臓で得た仮説をESあるいはカエルに使うことによって、若しくはその逆に、ESで得た仮説を腎臓に使うことによって、知識を深め、最終的には臓器の再生を目指したい。腎臓のように複雑な臓器の発生・再生は、研究者から敬遠される傾向にあったが、最近は徐々にScienceの土台にのりつつあり、基礎研究と臨床応用との境目が急速に接近している。このchallengingな課題に取り組むには、理学系と医学系の人間の共同作業が必要であると信じている。
幹細胞シグナル分子制御研究部の研究内容
 Annual Report 1998を見る
Annual Report 1999を見る
Annual Report 2000を見る
Annual Report 2002を見る
Annual Report 1998を見る
Annual Report 1999を見る
Annual Report 2000を見る
Annual Report 2002を見る
(This file is in Adobe Acrobat (PDF) format. If you do not have installed or need to upgrade to the latest version of Adobe Acrobat Reader, you can get a software from the Acrobat
web site)
研究室のホームページに戻る。
医科研のホームページに戻る。
最終更新日 2004年4月26日