1. ヒストンバリアントリン酸化による染色体分配制御
細胞分裂期における娘細胞への均等な染色体分配は、細胞あたりの染色体数を保つのみならず、安定的な染色体維持に必須の 事象である。細胞分裂期における均等な染色体分配は、染色体上に局在する様々なたんぱく質のリン酸化・脱リン酸化により 制御されている。The chromosomal passenger complex (CPC)は、Aurora Bキナーゼ、INCENP、Survivin、Borealinから なる複合体で、染色体分配の開始の制御に中心的な役割を果たしている。このうちAurora Bキナーゼは、細胞周期S期の後期に 活性化され、様々な基質をリン酸化することで精密な染色体分配をコントロールしている。このためAurora Bの活性は、細胞周期 の進行に伴い活性の強さ、及び場所が厳密に制御されている。Aurora BはS期後期においてINCENPとの結合のより部分的に活性化 され、ヒストンH3分子のセリン10をリン酸化し、HP1たんぱく質のヒストンH3分子からの遊離を促進する。その後、CPCは セントロメア領域に移行し、Aurora Bキナーゼのトレオニン232を自己リン酸化することで完全な活性化を受け、セントロメア 領域のたんぱく質をリン酸化することで、染色体分配が開始される。これまで、Aurora Bのセントロメアへの移行は、Bub1-H2A- Sugoshin経路と、Aurora B-Haspin-H3経路により制御されていることがわかっていたが、なぜセントロメアでAurora Bの完全 活性化が急激に、また局在して起こるのかについてはよくわかっていなかった。 我々は、ヒストンH2AのバリアントであるヒストンH2AXが染色体分配に重要な役割を果たしていることを見出した。これまで、 ヒストンH2AXはDNA二重鎖切断部位において、ATMキナーゼによりセリン139がリン酸化され、様々なDNA損傷応答たんぱく質 のリクルートに重要な役割を果たしていることが知られている。ヒストンH2AXノックアウト、ノックダウン細胞では染色体分配の 異常が顕著に見られた。またこの異常は、DNA二重鎖切断とは無関係で、Aurora B活性化異常によるものであることが分かった。 興味深いことに、Aurora BはヒストンH2AX分子のセリン121を特異的にリン酸化する事、さらにこのリン酸化がセントロメア 領域に局在することが明らかとなった(図1)。
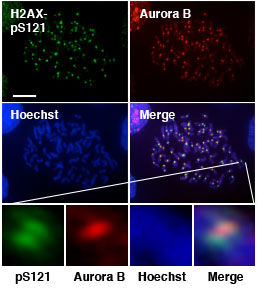
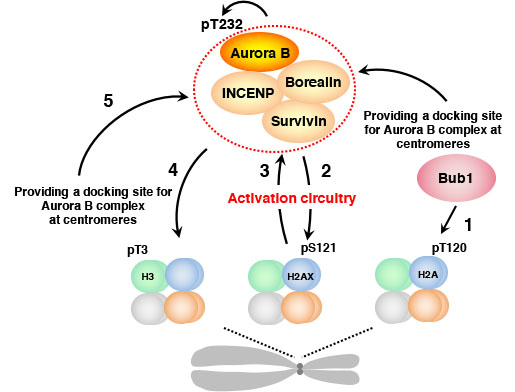
さらに、Aurora BによるヒストンH2AXのS121のリン酸化はAurora B分子のT232の自己リン酸化を促進するのに
重要な機能を果たしていることも分かった(図2)。
今後、この経路の異常が実際の発がん過程でどのような役割を果たしているのかについて解析を進めていく。